3. Хронофармакология пробиотиков при гастроэнтерите у телят
3.1. Влияние циркадианных и сезонных биологических ритмов на терапевтическую эффективность ветома 3 при гастроэнтерите у телят
Реакция на вещество во времени должна непременно варьировать в прямой зависимости от того, на какую фазу биоритма она приходится. По мнению Э.Б. Арушаняна (2000), это вынуждает к пересмотру существующих в экспериментальной практике и клинике количественной оценки фармакологического эффекта. В частности, резонно распространить на существующие сегодня экспериментальные подходы предложенный F. Halberg et al. (1962) критерий хронодесмы, под которым подразумеваются доверительные колебания какого-либо функционального показателя во времени.
В преимуществе такого приема перед существующими стационарными методами оценки убеждает тот факт, фармакологический ответ может быть выше или ниже средней величины, рассчитанной для стационарных условий, хотя он и не выходит за пределы физиологических колебаний той или иной функции. Напротив, реакция может ошибочно приниматься за адекватную, тогда как на самом деле оказывается ниже или выше нормальной величины, находясь при этом в зоне стационарной нормы (Арушанян Э.Б., 2000).
Согласно концепции F. Halberg и A. Reinberg (1967), при обычной терапии равномерно повторяемое одинаковое лекарственное воздействие каждый раз вызывает разный эффект вплоть до развития побочных реакций. При хронотерапии, когда препарат назначается в разных дозах на протяжении суток в зависимости от фаз биоритмов, чувствительности к нему, можно получить одинаковую желаемую реакцию без риска развития побочных эффектов.
Хронотерапия включает следующие методологические приемы: превентивные, имитационные, метод «навязывания ритма» (Комаров Ф.И., 1988) и метод учета хроночувствительности заинтересованных систем (функций) к фармакологическим веществам (Заславская Р.М., 1991, 1996).
Имитационный метод хронотерапии основывается на уже установленных закономерностях изменений концентрации определенных веществ в крови и тканях в соответствии с характерным для здорового индивидуума биоритмом (Заславская Р.М., 1991).
Метод «навязывания ритма» основан на использовании лекарственных и нелекарственных средств, для задания организму больного животного определенных ритмов, приближающихся к нормальным ритмам определенного вида.
Метод учета основан на выявлении хроночувствительности заинтересованных систем, в частности сердечно-сосудистой, к фармакологическим препаратам разных классов. Р.М. Заславская (1991, 1996), К.Ж. Ахметов (1993) применяли метод учета хроночувствительности заинтересованных систем в группах больных гипертонической болезнью. Используя данный метод, Ж.Ж. Кулкаева (1997) выявила времязависимые эффекты и суточный ритм чувствительности показателей гемодинамики к пропранополу, метопрололу, верапамилу, кордипину при лечении ишемической болезни сердца.
Изучению сезонных особенностей фармакологической чувствительности живых организмов посвящены работы А.П. Голикова и П.П. Голикова (1973); С.И. Рапопорта и соавт. (1987) и многих других. Так, по данным С.А. Лужновой и Н.Г. Урляповой (2004), степень влияния диаминодифинилсульфона на гематологические показатели мышей сопряжена с сезоном года. В работе О.Ю. Леденевой (2003) указывается на наличие сезонных особенностей фармакологического эффекта пробиотического препарата ветомгин при эндометрите у коров. В.М. Фещенко (2003) отмечает, что терапевтический эффект пробиотического препарата ветом 4 и низкоинтенсивного лазерного излучения при бронхопневмонии у поросят также зависит от сезона года.
Период биоритма любого функционального или структурного процесса регулируется ритмами его энергообеспечения. Присутствие в пространственной организации биологической системы градиентов протекающих в ней процессов должно учитываться при исследовании механизмов регуляции деятельности системы в качестве основы возникновения в ней различных реакций на влияние регулирующих факторов в зависимости от точки приложения их действия в градиенте, а также дает возможность избирательного пространственного градиентного воздействия на систему внешними факторами, в том числе лекарственными средствами (Комаров Ф.И., Рапопорт С.И., 2000).
Наличие существенных суточных колебаний эффективности терапевтического воздействия известно уже для многих классов лекарственных веществ, в частности для гипотензивных средств, противогистаминных, цитостатических, гормональных препаратов, наркотических средств, для многих из этих препаратов изучена хронотоксикология, а также сезонные колебания их действия (Зидермане А.А., 1988; Арушанян Э.Б., 1988, 2000).
Однако все вышеперечисленные данные в основном представлены из опытов на лабораторных животных, либо путем простого наблюдения за больными людьми, Сельскохозяйственные и домашние животные как объект биоритмологии, хронофармакологических исследований остаются «белым пятном» до сих пор.
С целью выявления зависимости терапевтической эффективности ветома 3 у телят, больных гастроэнтеритом, от хронобиологических факторов изучали особенности течения болезни при применении препарата в различное время суток, его влияние на морфологические и биохимические показатели периферической крови и интенсивность роста больных животных.
Для определения сезонных особенностей терапевтической эффективности ветома 3 провели две аналогичные серии опытов летом и осенью (табл. 2).
Исследования проводились на двухмесячных телятах черно-пестрой породы с характерными признаками гастроэнтерита. Диагноз был установлен на основании анамнеза и клинических признаков. Животных по принципу аналогов разделили на 5 опытных и контрольную группы животных по 5 телят в каждой. Животным опытных групп назначали ветом 3 в дозе 50 мг/кг массы дважды в сутки с водой в различное время: в 1-й группе – в 7 и 19 ч, 2-й – в 8 и 20 ч, 3-й – в 9 и 21 ч, 4-й – в 10 и 22 ч, 5-й – в 11 и 23 ч. Пробиотик назначали до полного клинического выздоровления. Телятам из контроля ветом 3 не применяли, животных лечили по схеме, традиционно применяемой в данном хозяйстве: в качестве антибактериального средства назначали гентамицина сульфат в дозе 2 мг/кг массы внутримышечно 2 раза в день. Кроме того, животным всех групп проводили регидратационную и восстановительную терапии (табл. 2).
Таблица 2. Схема опыта
| № гр. |
Тип Группы |
Препараты |
Путь введения |
Дозы и кратность применения |
Время назначения |
|
1 |
Опытные |
Ветом |
Внутрь |
мгкг рвдень |
7 и 19 ч. |
|
2 |
8 и 20 ч. |
||||
|
3 |
9 и 21 ч. |
||||
|
4 |
10 и 22 ч. |
||||
|
5 |
11 и 23 ч. |
||||
|
6 |
Контроль |
Гентамицина сульфат |
В/м |
2 мг/кг 2 р. в день |
Утро, вечер |
|
Для всех групп |
Глюкоза 40% Аскорб. к-та 5% NaCl изотонич. |
В/в |
15 мл 3 мл ad 50 мл |
Утро |
|
|
Тиамина хлорид 5% Пиридоксина гидрохлорид 5% Цианкобаламин |
П/к |
По 2 мл, чередовать |
Утро |
||
|
Тривит |
В/м |
5 мл |
Однократно |
||
Взятие крови проводили из яремной вены перед началом и по окончанию исследований.
Для изучения хронофармакологических особенностей влияния препарата на состояние неспецифической резистентности и обмена веществ, проводили гематологические и биохимические исследования крови.
Морфологические исследования крови включали определение уровня эритроцитов и лейкоцитов (меланжерным способом с использованием камеры Горяева), гемоглобина (по Сали), скорости оседания эритроцитов (по Панчекову). Кроме того, в мазках, окрашенных по Романовскому-Гимза, определяли лейкограмму (по Болотникову И.А., 1980).
Биохимические исследования крови включали определение уровня общего белка и белковых фракций (рефрактометрический метод), холестерина (ферментативный колориметрический метод), мочевины (фотометрический метод по реакции с диацетилмонооксимом), креатинина (реакция Яффе с последовательной фотометрией по Слоту), билирубина и его фракций (фотометрический модифицированный метод Йендрашека-Грофа), аланинаминотрансферазы и аспартатаминотрансферазы (фотометрический метод Райтмана-Френкеля), кальция (фотометрическое определение по реакции с арсеназо-III) и фосфора (фотометрическое определение по реакции с молибденовокислым аммонием), рассчитывали коэффициент де Ритиса и показатель фосфорно-кальциевого отношения (Комаров Ф.И. и соавт., 2001). Для проведения биохимических исследований использовали реактивы ЗАО «Вектор-Бест» (для определения мочевины – реактивы фирмы Lachema).
Временную организацию действия ветома 3 на показатели крови и живой массы животных изучали с помощью графически-параметрического метода анализа биологических ритмов (Романов Ю.А., 1979) и определяли следующие параметры: мезор – среднее значение показателей (фазы) за промежуток времени; акрофазу – момент крайнего отклонения фазы; активную (пассивную) фазы – значения фазы, лежащие выше (ниже) мезора; абсолютную амплитуду – разность между максимальным и минимальным значениями фазы; относительную амплитуду – отношение максимального к минимальному значению фазы; коэффициент синхронизации – отношение относительной амплитуды к промежутку времени между максимальным и минимальным значениями фазы (Комаров Ф.И., Рапопорт С.И., 2000).
Обработку данных проводили методами вариационной статистики с использованием программы «PGN-1.4». Достоверность результатов определяли с помощью критерия Стьюдента.
Установлено, что препарат обладает выраженным терапевтическим действием при гастроэнтерите. Улучшение состояния животных 1–5-й опытных групп, как в летний, так и в осенний периоды, отмечалось уже на 2-3-и сутки после начала лечения. У животных появлялся аппетит, нормализовались температура, пульс и частота дыхания; дефекация отмечалась реже, кал приобретал кашицеобразную консистенцию, уменьшалось количество не переваренных остатков корма. В то время как у животных контрольной группы заметных изменений в течение болезни не отмечали.
Температура тела у телят 1–5-й опытных групп после выздоровления в летний период была ниже по сравнению с контролем на 2,31; 1,29; 1,03; 1,54; 1,03 %, а по сравнению с исходными данными – на 2,56; 1,79; 1,53; 1,29; 1,03% соответственно, тогда как у животных из контроля температура тела по сравнению с исходными данными не изменилась. Необходимо отметить, что, если в 1-й и 2-й опытных группах отмечается плавное стабильной снижение температуры за весь период лечения, то в контрольной группе отмечается даже некоторый подъем температуры в первые 3-е суток лечения. В 3–5-й опытных группах также отмечалось снижение температуры, но более медленное с редкими периодами подъема (рис. 5).
В активной фазе (в данном случае ниже мезора, т.к. изначально вследствие воспалительной реакции, обусловленной гастроэнтеритом, температура была повышена) влияния ветома 3 на уровень температуры у телят в летний период находились показатели 1-й и 4-й опытных групп, которые были ниже мезора (38,34±0,15 °С) на 0,9 (P<0,05) и 0,3 %, в пассивной фазе – показатели 2, 3 и 5-й опытных групп, которые превышали мезор на 0,2; 0,4 и 0,4 % соответственно. Продолжительность активной фазы была в 4 раза короче пассивной фазы. Абсолютная амплитуда показателей температуры телят составила 0,5 °С, относительная амплитуда – 1,01 ед. Коэффициент синхронизации влияния ветома 3 на уровень температуры телят составил 0,51 ед/ч. Акрофаза (в данном случае минимальное значение) влияния ветома 3 на уровень температуры телят приходилась на 7 и 19 ч, что соответствует показателям телят 1-й опытной группы (рис. 5).
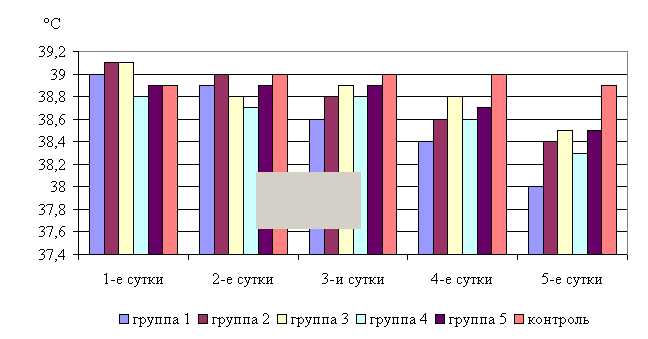
Рис. 5. Динамика показателей температуры телят в летний период
Температура тела у телят 1–5-й опытных групп после выздоровления осенью была ниже по сравнению с контролем на 1,54 (P<0,1); 1,8 (P<0,05); 1,03; 1,29 (P<0,01); 0,51 % соответственно. По сравнению с исходными данными температура тела у телят 1–5-й опытных и контрольной групп была ниже на 2,29; 2,3; 1,03; 2,29; 0,77 и 0,26 % соответственно. Необходимо отметить, что если во 2-й и 4-й опытных группах отмечается плавное и стабильное снижение температуры за весь период лечения, то в контрольной группе отмечается даже некоторый подъем температуры на 3-4-е сутки лечения. В 1, 3 и 5-й опытных группах также отмечалось снижение температуры, но более медленное с редкими периодами подъема (рис. 7).
Примечание: здесь и далее АФ – активная фаза, Акрф – акрофаза
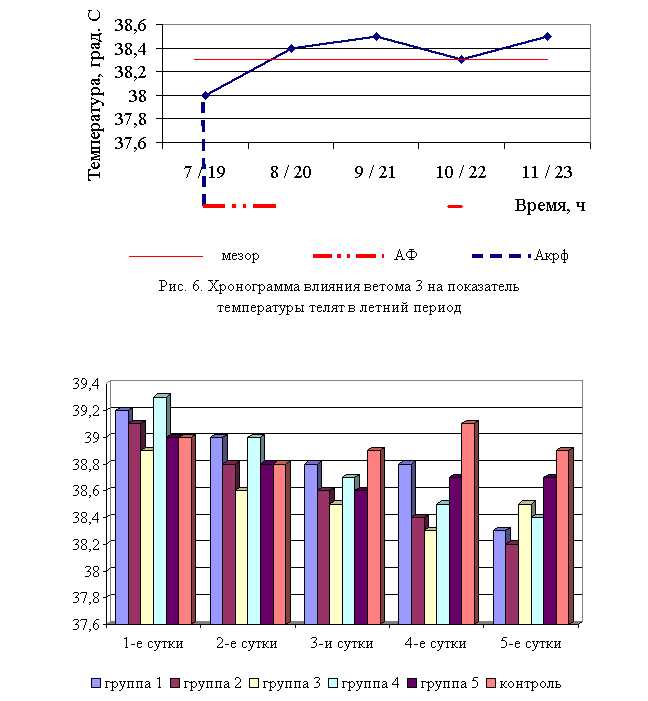
Рис. 7. Динамика показателей температуры телят в осенний период
В активной фазе (в данном случае ниже мезора, т. к. изначально вследствие воспалительной реакции, обусловленной гастроэнтеритом, температура была повышена) влияния ветома 3 на уровень температуры телят в осенний период находились показатели 1, 2 и 4-й опытных групп, которые были ниже мезора (38,42±0,08 °С) на 0,3, 0,6 (P<0,1) и 0,1 %, в пассивной фазе – показатели 3 и 5-й опытных групп, которые превышали мезор на 0,2 и 0,7 % соответственно. Продолжительность активной фазы была на 28,6 % длиннее пассивной фазы. Абсолютная амплитуда показателей температуры телят составила 0,5 °С, относительная амплитуда – 1,01 ед. Коэффициент синхронизации влияния ветома 3 на уровень температуры телят составил 0,34 ед/ч. Акрофаза (в данном случае минимальное значение) влияния ветома 3 на уровень температуры телят приходилась на 8 и 20 ч, что соответствует показателям телят 2-й опытной группы (рис. 8).
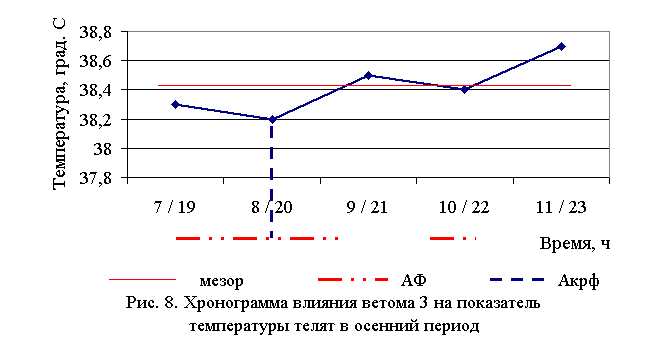
Таким образом, под влиянием ветома 3 происходило стабильное снижение температуры у подопытных телят. Выраженность фармакологического эффекта пробиотика находилась в зависимости от времени применения препарата: коэффициент синхронизации хронограмм в летний и осенний периоды составил 0,51 и 0,34 ед./ч. Оптимальное снижение температуры при применении препарата в летний период отмечено в 7 и 19 ч, осенью – в 8 и 20 ч, при этом показатели температуры были достоверно ниже мезора на 0,9 и 0,6 % соответственно.
При проведении опыта отмечены также изменения частоты пульса у подопытных телят.
У телят 1, 2 и 4-й опытных групп после выздоровления в летний период частота пульса была недостоверно ниже по сравнению с контролем на 1,56 % соответственно, тогда как у животных 3-й и 5-й опытных групп данный показатель находился на уровне контроля. По сравнению с исходными данными частота пульса у телят 1-й опытной и контрольной групп недостоверно снизилась на 3,1 и 1,5 % соответственно, тогда как у животных 2-й и 5-й опытных групп – увеличилась на 1,6 и 3,2 %, а у аналогов 3-й и 5-й опытных групп – не изменилась по сравнению с исходными данными (рис. 9).
Анализ хронобиологических особенностей влияния ветома 3 на частоту пульса в летний период не позволяет сделать однозначных выводов.
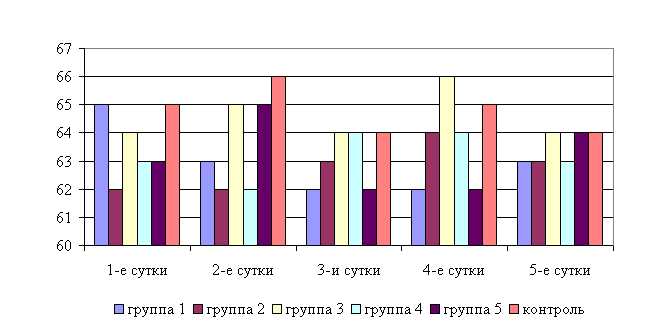
Рис. 9. Динамика частоты пульса у подопытных телят в летний период
У телят 1, 2, 3 и 4-й опытных групп после выздоровления в осенний период частота пульса была недостоверно ниже по сравнению с контролем на 1,56; 1,56; 3,12 и 1,56 % соответственно, тогда как у животных 5-й опытной группы данный показатель находился на уровне контроля. По сравнению с исходными данными частота пульса у телят 1, 2, 3 и 4-й опытных групп недостоверно снизилась на 3,08; 3,08; 1,59; 1,56 % соответственно, тогда как у животных 5-й опытной группы – увеличилась на 1,59 %, а у аналогов из контроля – не изменилась по сравнению с исходными данными. Необходимо отметить, что в течение опыта стабильно понижалась частота пульса только у телят 2-й опытной группы, тогда как у животных 1, 3, 4, 5-й опытных и контрольной групп снижение частоты пульса чередовалось с периодами подъема (рис. 10).
Анализ хронобиологических особенностей влияния ветома 3 на частоту пульса в осенний период также не позволяет сделать однозначных выводов.
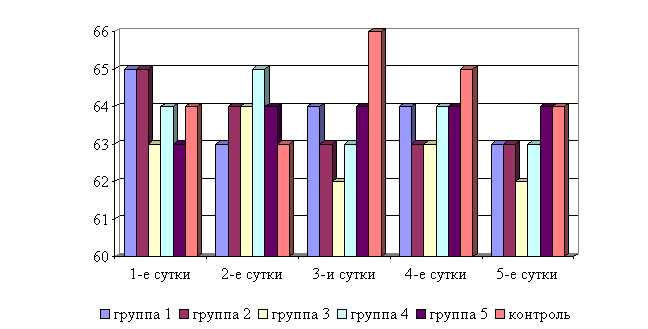
Рис. 10. Динамика частоты пульса у подопытных телят в осенний период
При проведении опыта отмечены также изменения частоты дыхательных движений у подопытных телят.
У телят 1, 2 и 3-й опытных групп после выздоровления в летний период частота дыхательных движений была недостоверно ниже по сравнению с контролем на 5,6; 5,6 и 11,1 % соответственно, тогда как у животных 4-й и 5-й опытной группы данный показатель находился на уровне контроля. По сравнению с исходными данными, частота дыхательных движений у телят 1–5-й опытных групп недостоверно снизилась на 5,6; 10,5; 15,8; 10,0; 5,3 % соответственно, тогда как у животных контрольной группы данный показатель не изменился по сравнению с исходными данными (рис. 11).
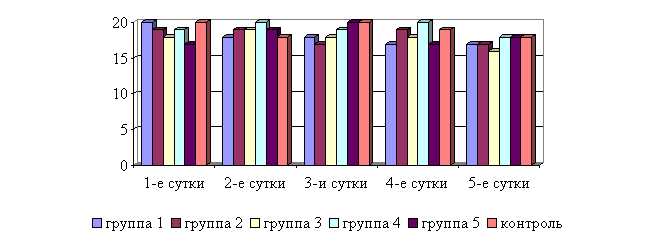
Рис. 11. Динамика частоты дыхательных движений у подопытных телят в летний период
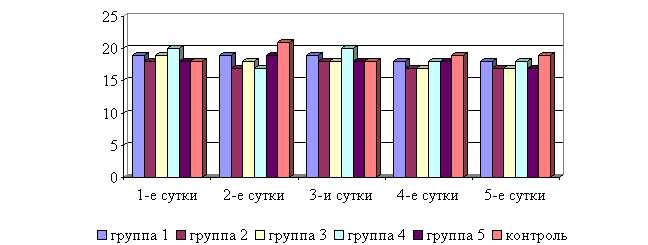
Рис. 12. Динамика частоты дыхательных движений у подопытных телят в осенний период
У животных 1–5-й опытных групп после выздоровления в осенний период частота дыхательных движений была недостоверно ниже по сравнению с контролем на 5,3; 10,5; 10,5; 5,3; 10,5 % соответственно. По сравнению с исходными данными частота дыхательных движений у телят 1–5-й опытных групп недостоверно снизилась на 5,3; 5,6; 10,5; 10,0; 5,6 % соответственно, тогда как у животных контрольной группы – увеличилась на 5,6 % (рис.12).
Таким образом, при применении ветома 3 как в летний, так и осенний периоды, отмечена тенденция к снижению частоты пульса и дыхательных движений, однако, анализ хронобиологических особенностей влияния препарата на данные показатели не позволяет сделать однозначных выводов.
При применении ветома 3 отмечено снижение продолжительности лечения телят опытных групп.
В летний период продолжительность лечения телят 1–5-й опытных групп была достоверно ниже по сравнению с аналогами из контроля на 52,25 (P<0,01); 40,93 (P<0,05); 29,6; 29,6; 22,65 (P<0,1) % соответственно (табл. 10).
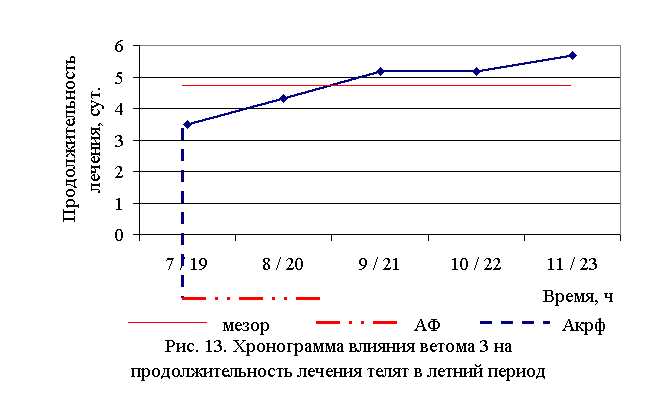
Наименьшая продолжительность лечения телят была отмечена в 1-й опытной группе, где уже на 3-и сутки опыта половина животных была клинически здорова, и средняя продолжительность лечения составила 3,5±0,42 суток. В контрольной группе показатель продолжительности лечения составил 7,33±0,61 суток и на 3-и сутки лечения не было ни одного выздоровевшего животного (табл. 10).
В активной фазе (в данном случае ниже мезора) влияния ветома 3 на продолжительность лечения телят в летний период находились показатели 1-й и 2-й опытных групп, которые были ниже мезора ((4,76±0,68) суток) на 26,5 и 9,0 %, в пассивной фазе – показатели 3, 4 и 5-й опытных групп, которые превышали мезор на 8,4; 8,4 и 19,1 % соответственно. Продолжительность активной фазы была на 40 % короче пассивной фазы. Абсолютная амплитуда показателей продолжительности лечения телят составила 2,17 суток, относительная амплитуда – 1,62 ед. Коэффициент синхронизации влияния ветома 3 на продолжительность лечения телят составил 0,41 ед./ч. Акрофаза (минимальное значение) влияния ветома 3 на продолжительность лечения телят летом приходилась на 7 и 19 ч, что соответствует показателям телят 1-й опытной группы (рис. 13).
Таблица 10
Продолжительность лечения телят в летний период
|
№ группы |
Время назначения ветома 3 |
Количество животных, выздоровевших на3 / 5 сутки опыта |
Средняя продолжительность лечения в группе, сут. |
|
1-я |
7 и 19 ч. |
3 / 2 |
3,5±0,42*** |
|
2-я |
8 и 20 ч. |
2 / 3 |
4,33±0,49** |
|
3-я |
9 и 21 ч. |
1 / 3 |
5,16±0,54* |
|
4-я |
10 и 22 ч. |
1 / 2 |
5,16±0,60* |
|
5-я |
11 и 23 ч. |
0 / 3 |
5,67±0,49* |
|
Контроль |
пробиотики не назначали |
0 / 1 |
7,33±0,61 |
В осенний период продолжительность лечения телят 1–5-й опытных групп была достоверно ниже, по сравнению с аналогами из контроля на 37,24 (P<0,05); 41,84 (P<0,01); 32,64 (P<0,05); 25,66 (P<0,1); 23,29 (P<0,1) % соответственно.
Наименьшая продолжительность лечения была отмечена во 2-й группе и составила 4,17±0,4 суток, тогда как в контрольной группе продолжительность лечения составила 7,17±0,4 суток (P<0,05). В 1, 2 и 3-й опытных группах уже на 3-и сутки опыта 2 из 5-ти животных были клинически здоровы, в контрольной группе на 3-и сутки лечения не было ни одного выздоровевшего животного (табл. 11) Таблица 11
Продолжительность лечения телят в осенний период
|
№ группы |
Время назначения ветома 3 |
Количество животных, выздоровевших на3 / 5 день лечения |
Средняя продолжительность лечения в группе |
|
1-я |
7 и 19 ч. |
2 / 3 |
4,5±0,50** |
|
2-я |
8 и 20 ч. |
2 / 4 |
4,17±0,40*** |
|
3-я |
9 и 21 ч. |
2 / 2 |
4,83±0,65** |
|
4-я |
10 и 22 ч. |
1 / 2 |
5,33±0,56* |
|
5-я |
11 и 23 ч. |
1 / 2 |
5,5±0,62* |
|
контроль |
Пробиотики не назначали |
0 / 0 |
7,17±0,40 |
В активной фазе (в данном случае ниже мезора) влияния ветома 3 на продолжительность лечения телят в осенний период находились показатели 1, 2 и 3-й опытных групп, которые были ниже мезора ((4,87±0,25) суток) на 7,6; 14,4 и 0,8 %, в пассивной фазе – показатели 4 и 5-й опытных групп, которые превышали мезор на 9,4 и 12,9 % соответственно. Продолжительность активной и пассивной фаз были равными. Абсолютная амплитуда показателей продолжительности лечения телят составила 1,33 суток, относительная амплитуда – 1,32 ед. Коэффициент синхронизации влияния ветома 3 на продолжительность лечения телят составил 0,44 ед./ч. Акрофаза (минимальное значение) влияния ветома 3 на продолжительность лечения телят осенью приходилась на 8 и 20 ч, что соответствует показателям телят 2-й опытной группы (рис. 14).
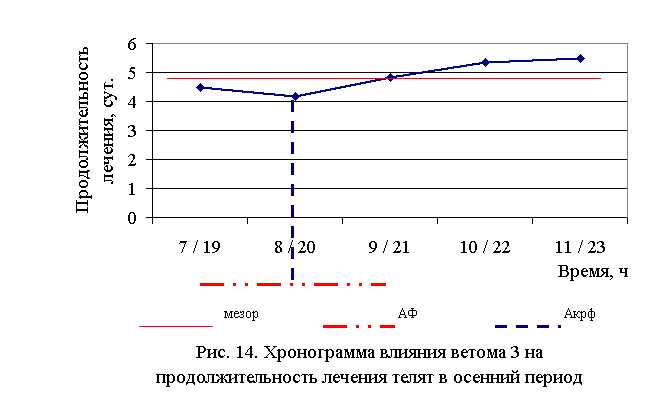
Таким образом, ветом 3 обладает выраженным терапевтическим эффектом при гастроэнтерите у телят, который находится в прямой зависимости от времени введения препарата и сезона года – коэффициент синхронизации влияния пробиотика на продолжительность лечения в летний и осенний периоды составлял 0,41 и 0,44 ед./ч соответственно. Оптимальный терапевтический эффект получен при применении препарата в 7 и 19 ч в летний период, и в 8 и 20 ч – осенью, при этом продолжительность лечения телят была ниже мезора на 26,5 и 14,4 % соответственно. Ветом 3 при применении в указанные временные рамки вызывал стабильное снижение температуры, причем акрофаза показателя была ниже мезора на 0,9 и 0,6 % в летний и осенний периоды соответственно.
Полученные результаты согласуются с данными ряда авторов о зависимости физиологических показателей живых организмов от фазы биологического ритма. Такие ритмы называют физиологическими. Влияние биологических ритмов на физиологические показатели закреплено эволюционно и основано на различной восприимчивости нейрогуморальных структур к внешним воздействиям (свет, температура окружающей среды и т.д.) в разные периоды суток. Соответвенно, терапевтическое воздействие на живой организм в различные периоды активности физиологических систем вызывает различный эффект. По нашим данным, оптимальное влияние ветома 3 на физиологические показатели телят, больных гастроэнтеритом, различно в летний и осенний периоды, что в свою очередь согласуется с концепцией сезонной организации биологических систем (сезонные биологические ритмы).
3.2. Хронобиологические особенности действия ветома 3 на морфологические показатели крови у телят после выздоровления при гастроэнтерите
В результате проведенных исследований установлено, что до применения ветома 3 количество эритроцитов, лейкоцитов и гемоглобина у телят опытных и контрольной групп не имело достоверных различий и находилось в пределах физиологических норм.
Анализ лабораторных исследований свидетельствует о повышении уровня эритроцитов в крови телят опытных групп после проведения лечения в летний и осенний периоды (табл. 12).
Количество эритроцитов у телят 1–5-й опытных групп после выздоровления в летний период было выше, по сравнению с аналогами из контроля на 8,24; 0,33; 1,81; 7,99 (P<0,1); 4,12 % соответственно. По сравнению с исходными данными уровень эритроцитов повысился у животных 1–5-й опытных групп на 3,63; 1,42; 1,23; 2,34; 0,24 % соответственно, а у аналогов из контроля – снизился на 1,62 % (табл. 12).
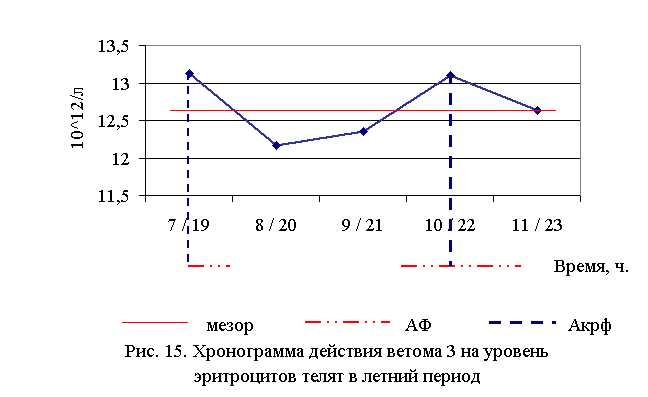
В активной фазе влияния ветома 3 на уровень эритроцитов у телят в летний период находились показатели 1-й и 4-й опытных групп, которые превышали мезор ((12,68±0,19), 1012/л) на 3,5 и 3,3 % соответственно, в пассивной фазе – показатели 2, 3 и 5-й опытных групп, которые были ниже мезора на 4,0; 2,6 и 0,4% соответственно. Продолжительность активной и пассивной фаз были равными. Абсолютная амплитуда была равной 0,96 1012/л, относительная амплитуда – 1,08 ед. Коэффициент синхронизации влияния ветома 3 на уровень эритроцитов у телят составил 1,08 ед./ч. Акрофаза влияния ветома 3 на количество эритроцитов у телят приходилась на 7 и 19, 10 и 22 ч, что соответствует показателям телят 1-й и 4-й опытных групп (рис. 15).
Количество эритроцитов у телят 1–5-й опытных групп после выздоровления в осенний период было выше по сравнению с аналогами из контроля на 11,11 (P<0,05); 11,97 (P<0,05); 9,15 (P<0,05); 1,71; 2,31 %, а по сравнению с исходными данными – на 3,42; 10,74 (P<0,05); 4,93; 1,45; 3,19 % соответственно. Тогда как у аналогов из контроля уровень эритроцитов по сравнению с исходными данными снизился на 1,62 % (табл. 12).
Таблица 12. Динамика уровня эритроцитов у телят, 1012/л
| Группа |
Лето |
Осень |
||
|
До лечения |
После выздоровления |
До лечения |
После выздоровления |
|
|
1-я опытная |
12,67±0,37 Cv=5,08 |
13,13±0,33 Cv=4,40 |
12,57±0,44 Cv=6,03 |
13,0±0,42** Cv=5,55 |
|
2-я опытная |
12,0±0,7 Cv=10,1 |
12,17±0,34 Cv=4,82 |
11,83±0,09 Cv=1,29 |
13,1±0,4** Cv=5,18 |
|
3-я опытная |
12,2±0,53 Cv=7,51 |
12,35±0,41 Cv=5,96 |
12,17±0,44 Cv=6,28 |
12,77±0,29** Cv=3,86 |
|
4-я опытная |
12,8±1,1 Cv=14,91 |
13,1±0,24* Cv=3,17 |
11,73±0,44 Cv=6,51 |
11,9±0,44 Cv=6,34 |
|
5-я опытная |
12,6±0,5 Cv=6,92 |
12,63±0,43 Cv=5,85 |
11,6±0,57 Cv=8,49 |
11,97±0,29 Cv=4,21 |
|
контроль |
12,33±0,29 Cv=4,08 |
12,13±0,43 Cv=6,24 |
11,37±0,29 Cv=4,43 |
11,7±0,17 Cv=2,5 |
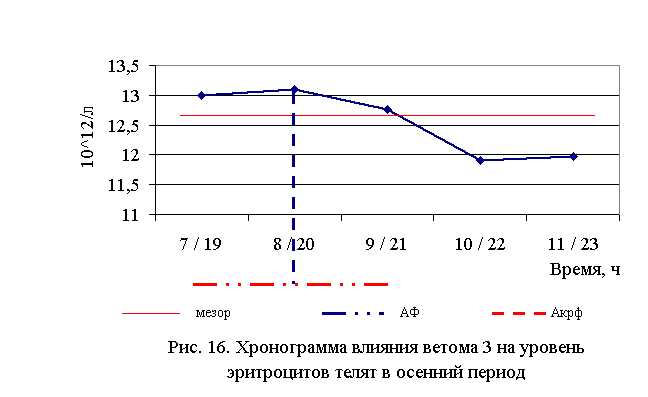
В активной фазе влияния ветома 3 на уровень эритроцитов телят в осенний период находились показатели 1, 2 и 3-й опытных групп, которые превышали мезор (12,68±0,19, 1012/л) на 2,5, 3,3 и 0,7% соответственно, в пассивной фазе – показатели 4-й и 5-й опытных групп, которые были ниже мезора на 6,2 и 5,6% соответственно. Продолжительность активной фазы была больше продолжительности пассивной на 40%. Абсолютная амплитуда была равной 1,2, 1012/л, относительная амплитуда – 1,1 ед. Коэффициент синхронизации влияния ветома 3 на уровень эритроцитов у телят составил 0,55 ед./ч. Акрофаза влияния ветома 3 на количество эритроцитов у телят приходилась на 8 и 20 ч, что соответствует показателям телят 2-й опытной группы (рис. 16).
Таким образом, ветом 3 повышает уровень эритроцитов у телят в пределах физиологических норм. Выраженность влияния препарата на данный показатель находится в зависимости от времени введения пробиотика и сезона года. Оптимальный эффект получен при применении ветома 3 в 7 и 19, и 10 и 22 ч – летом, и в 8 и 20 ч – осенью. При применении препарата в эти временные рамки уровень эритроцитов был выше мезора на 3,5 и 3,3 % в летний и осенний периоды соответственно. Под влиянием ветома 3 содержание гемоглобина у телят опытных групп также изменялось (табл. 13). Содержание гемоглобина у телят 1–5-й опытных групп после выздоровления в летний период было достоверно выше по сравнению с аналогами из контроля на 9,5; 6,4; 5,2; 6,1; 6,2 % соответственно. По сравнению с исходными данными уровень гемоглобина повысился у телят 1–5-й опытных и контрольной групп на 11,8; 9,6; 7,8; 8,2; 7,7 и 2,3 % соответственно (табл. 13)
Таблица 13. Динамика содержания гемоглобина у телят, г/л
Группа |
Лето |
Осень |
||
|
До |
После выздоровления |
До |
После выздоровления |
|
|
1-я опытная |
121,6±1,03 |
136,0±1,82 # |
121,2±2,42 |
125,6+1,21* |
|
2-я опытная |
120,6±1,08 |
132,2±2,01*** |
124,2±1,42 |
127,6±0,93*** |
|
3-я опытная |
121,2±1,69 |
130,6±1,25*** |
122,0±1,82 |
124,4±0,6 |
|
4-я опытная |
121,8±1,5 |
131,8±1,52*** |
122,2±1,59 |
125,0±0,77* |
|
5-я опытная |
122,6±1,66 |
132,0±1,0 # |
121,8±1,71 |
126,6±2,99 |
|
контроль |
121,4±1,03 |
124,2±0,74 |
122,2±1,43 |
122,8±0,66 |
Примечание: здесь и далее # - P<0,001
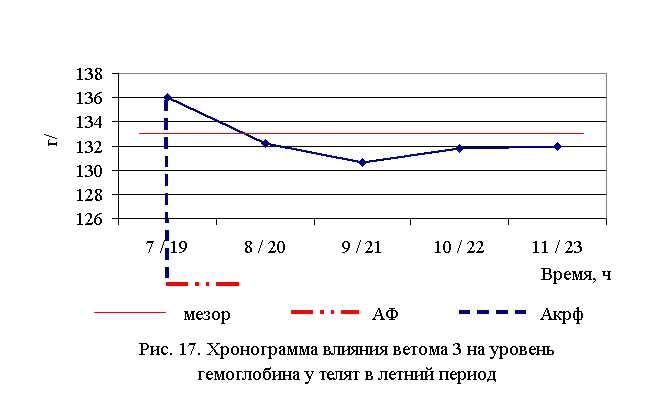
В активной фазе влияния ветома 3 на уровень гемоглобина телят в летний период находились показатели 1-й опытной группы, которые превышали мезор ((132,52±0,91) г/л) на 2,6 % соответственно, в пассивной фазе – показатели 2–5-й опытных групп, которые были ниже мезора на 0,2; 1,4; 0,5; 0,4 % соответственно. Продолжительность активной фазы была короче пассивной на 76,9 %. Абсолютная амплитуда была равной 5,4 г/л, относительная амплитуда – 1,04 ед. Коэффициент синхронизации влияния ветома 3 на уровень гемоглобина у телят в летний период составил 0,52 ед./ч. Акрофаза влияния ветома 3 на содержание гемоглобина у телят приходилась на 7 и 19 ч, что соответствует показателям телят 1-й опытной группы (рис. 17).
Содержание гемоглобина у телят 1–5-й опытных групп после выздоровления в осенний период было выше по сравнению с аналогами из контроля на 2,3 (P<0,1); 3,9 (P<0,01); 1,3; 1,8 (P<0,1); 3,1 % соответственно. По сравнению с исходными данными уровень гемоглобина повысился у телят 1–5-й опытных и контрольной групп на 3,6; 2,7; 2,0; 2,3; 3,9 и 0,5 % соответственно (табл. 14).
В активной фазе влияния ветома 3 на уровень гемоглобина телят в осенний период находились показатели 2-й и 5-й опытной группы, которые превышали мезор ((132,52±0,91) г/л) на 1,4 и 0,6 % соответственно, в пассивной фазе – показатели 1, 3 и 4-й опытных групп, которые были ниже мезора на 0,2; 0,8 и 0,7 % соответственно. Продолжительность активной фазы была длиннее пассивной на 28,6 %. Абсолютная амплитуда была равной 3,2 г/л, относительная амплитуда – 1,03 ед. Коэффициент синхронизации влияния ветома 3 на уровень гемоглобина у телят в летний период составил 1,03 ед./ч. Акрофаза влияния ветома 3 на содержание гемоглобина у телят приходилась на 8 и 20 ч, что соответствует показателям телят 2-й опытной группы (рис. 18).
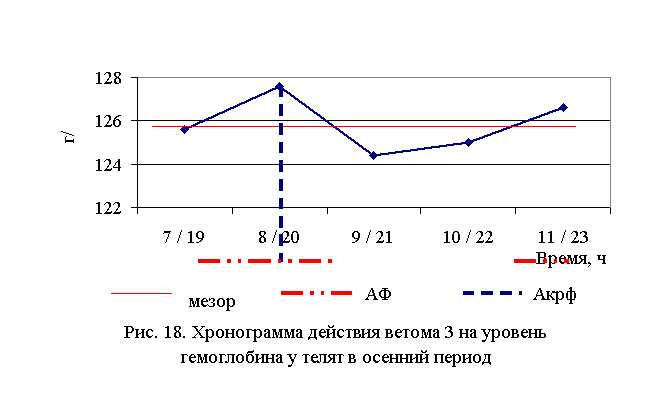
Таким образом, ветом 3 повышает уровень гемоглобина у телят в пределах физиологических норм. Выраженность влияния препарата на данный показатель находится в зависимости от времени введения и сезона. Оптимальный эффект получен при применении пробиотика в 7 и 19 ч – летом и в 8 и 20 ч – осенью, при применении препарата в эти временные рамки уровень эритроцитов был выше мезора на 2,6 и 1,4 % соответственно.
Под влиянием ветома 3 уровень лейкоцитов в крови телят опытных групп также изменялся (табл. 14).
Количество лейкоцитов у телят 1, 4 и 5-й опытных групп после выздоровления в летний период было выше по сравнению с аналогами из контроля на 5,39; 1,13; 1,13 %, а у животных 2-й и 3-й опытных групп – ниже на 4,26 и 3,22 % соответственно. По сравнению с исходными данными уровень лейкоцитов повысился у телят 1–5-й опытных групп на 14,91 (P<0,001); 8,48; 2,21; 3,88; 9,02 % соответственно, а у аналогов из контроля – снизился на 1,11 % (табл. 14)
Таблица 14. Динамика уровня лейкоцитов у телят, 109/л
Группа |
Лето |
Осень |
||
|
До |
После выздоровления |
До |
После выздоровления |
|
|
1-я опытная |
11,4±0,15 |
13,1±0,12 |
12,2±0,08 |
13,17±0,07** |
|
2-я опытная |
10,97±0,87 |
11,9±0,42 |
11,93±0,47 |
13,0±0,15* |
|
3-я опытная |
11,77±0,27 |
12,03±0,06 |
12,5±0,69 |
12,63±0,63 |
|
4-я опытная |
12,1±0,11 |
12,57±0,24 |
12,7±0,8 |
12,77±0,35 |
|
5-я опытная |
11,53±1,33 |
12,57±0,67 |
11,77±0,77 |
12,17±0,57 |
|
контроль |
12,57±0,73 |
12,43±0,75 |
11,9±0,31 |
12,17±0,35 |
В активной фазе влияния ветома 3 на уровень лейкоцитов у телят в летний период находились показатели 1, 4 и 5-й опытных групп, которые превышали мезор ((12,43±0,38) 109/л) на 5,4; 1,1 и 1,1 % соответственно, в пассивной фазе – показатели 2-й и 3-й опытных групп, которые были ниже мезора на 4,3 и 3,2 % соответственно. Продолжительность активной фазы была больше пассивной на 28,6 %. Абсолютная амплитуда была равной 1,2* 109/л, относительная амплитуда – 1,1 ед. Коэффициент синхронизации влияния ветома 3 на уровень лейкоцитов у телят составил 1,1 ед./ч. Акрофаза влияния ветома 3 на количество лейкоцитов у телят приходилась на 7 и 19 ч, что соответствует показателям телят 1-й опытной группы (рис. 19).
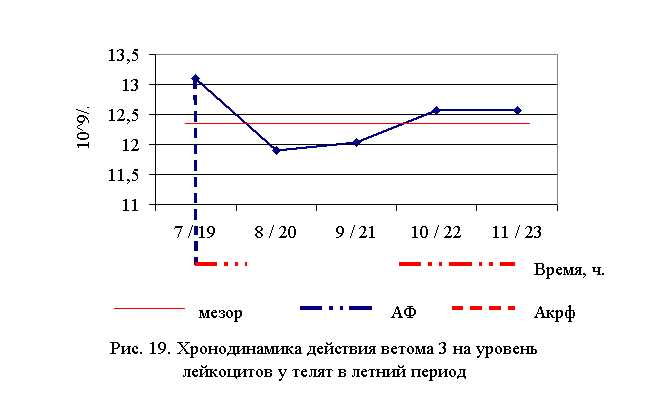
Количество лейкоцитов у телят 1–4-й опытных групп после выздоровления в осенний период было выше по сравнению с аналогами из контроля на 8,22 (P<0,05), 6,82 (P<0,1), 3,78, 4,93%, а у животных 5-й опытных групп – на уровне показателей контрольной группы. По сравнению с исходными данными уровень лейкоцитов повысился у телят 1–5-й опытных и контрольной групп на 7,95 (P<0,001), 8,97 (P<0,1), 1,04, 0,55, 3,40 и 2,27% соответственно (табл. 14).
В активной фазе влияния ветома 3 на уровень лейкоцитов телят в осенний период находились показатели 1, 2 и 4-й опытных групп, которые превышали мезор (12,75±0,17, 109/л) на 3,3 (P<0,1), 2,0 и 0,2% соответственно, в пассивной фазе – показатели 3-й и 5-й опытных групп, которые были ниже мезора на 0,9 и 4,5% соответственно. Продолжительность активной фазы была больше пассивной на 8,7%. Абсолютная амплитуда была равной 1,0, 109/л, относительная амплитуда – 1,1 ед. Коэффициент синхронизации влияния ветома 3 на уровень лейкоцитов
у телят составил 0,3 ед./ч. Акрофаза влияния ветома 3 на количество лейкоцитов у телят приходилась на 7 и 19 ч, что соответствует показателям телят 1-й опытной группы (рис. 20).
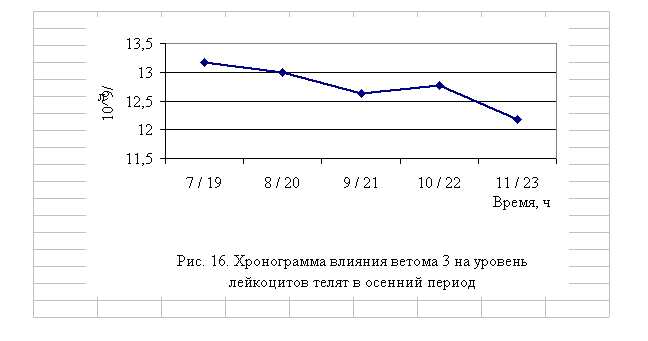
Таким образом, ветом 3 повышает уровень лейкоцитов у телят в пределах физиологических норм. Выраженность влияния препарата на данный показатель находится в зависимости от времени введения препарата и сезона года. Оптимальный эффект получен при применении пробиотика в 7 и 19 ч летом и осенью, при применении препарата в эти временные рамки уровень эритроцитов был выше мезора на 5,4 и 2,0 % соответственно.
Нами установлены изменения в показателе скорости оседания эритроцитов. Первоначально этот показатель по всем группам составлял в среднем в летний период 2,5 мм/ч, в осенний – 2,25 мм/ч. После выздоровления он значительно уменьшился и составил в летний период – 1,0 мм/ч в 1, 2 и 4-й группах, 1,5 мм/ч – в 3-й и 5-й группах, в контрольной группе данный показатель остался без изменений и составил 2 – 2,5 мм/ч у всех животных, в осенний период – у телят 1, 2 и 5-й групп – 1,0 мм/ч, 3-й и 4-й групп – 1,5 мм/ч, у телят из контроля данный показатель практически не изменился по сравнению с исходными данными.
При применении ветома 3 отмечены изменения в процентном соотношении лейкоцитарных клеток у телят.
Уровень лимфоцитов до применения препарата был ниже физиологической нормы (табл. 15).
Процентное содержание лимфоцитов после выздоровления в летний период у телят 1–5 опытных групп было выше по сравнению с аналогичным показателем животных из контроля на 6,7 (P<0,1); 5,3 (P<0,05); 4,6; 2,8; 3,5%. По сравнению с исходными данными содержание лимфоцитов увеличилось у телят 1–5-й опытных и контрольной групп на 5,2 (P<0,1); 8,3 (P<0,05); 1,7; 3,2; 3,5 и 1,8 % соответственно (табл. 15-16).
В активной фазе влияния ветома 3 на уровень лимфоцитов в летний период находились показатели 1-й и 2-й опытных групп, которые превышали мезор (59,4±0,38) % на 2,0 и 0,7 %, в пассивной фазе находились показатели 4-й и 5-й опытных групп, которые были ниже мезора на 1,7 и 1,0 % соответственно, уровень лимфоцитов 3-й опытной группы соответствовал мезору (рис. 19).
Таблица 15. Лейкоцитарный профиль телят до лечения в летний период, %
| Показатель |
Опытные группы |
Контроль |
||||
|
1-я |
2-я |
3-я |
4-я |
5-я |
||
|
Э |
0,4±0,24 Сv=136,93 |
0,8±0,2 Cv=55,9 |
0 |
0,4±0,24 Cv=136,93 |
0,6±0,4 Cv=149,07 |
1,2±0,2 Cv=37,27 |
|
Б |
0 |
0 |
0 |
0 |
0 |
0 |
|
Ю |
0,8±0,2 Cv=55,9 |
1,2±0,37 Cv=69,72 |
0,8±0,37 Cv=104,58 |
0,8±0,49 Cv=136,93 |
1,0±0,55 Cv=122,47 |
0,4±0,4 Cv=223,61 |
|
П |
6,2±0,2 Cv=7,21 |
7,2±0,37 Cv=11,62 |
6,6±0,51 Cv=17,28 |
5,8±0,37 Cv=14,43 |
6,2±0,58 Cv=21,03 |
7,4±0,24 Cv=7,4 |
|
С |
32,4±0,6 Cv=4,25 |
32,4±0,75 Cv=5,16 |
31,2±1,24 Cv=8,89 |
32,8±0,2 Cv=1,36 |
31,4±0,98 Cv=6,98 |
31,0±0,89 Cv=6,45 |
|
Л |
57,6±0,6 Cv=2,29 |
55,2±1,71 Cv=6,95 |
58,4±2,11 Cv=8,09 |
56,6±1,36 Cv=5,39 |
56,8±1,53 Cv=6,02 |
55,8±2,13 Cv=8,54 |
|
М |
2,6±0,6 Cv=51,6 |
3,2±0,37 Cv=26,15 |
3,0±0,77 Cv=57,74 |
3,6±0,6 Cv=37,27 |
4,0±1,06 Cv=58,63 |
4,2±0,92 Cv=48,8 |
Продолжительность активной и пассивной фаз при проведении опыта в летний период была равной. Абсолютная амплитуда показателей уровня лимфоцитов составила 2,2%, относительная амплитуда – 1,04 ед. Коэффициент синхронизации влияния ветома 3 на содержание лимфоцитов у телят в летний период составил 0,35 ед./ч. Акрофаза влияния ветома 3 на уровень лимфоцитов у телят приходилась на 7 и 19 ч, что соответствует показателям телят 1-й опытной группы (рис. 21).
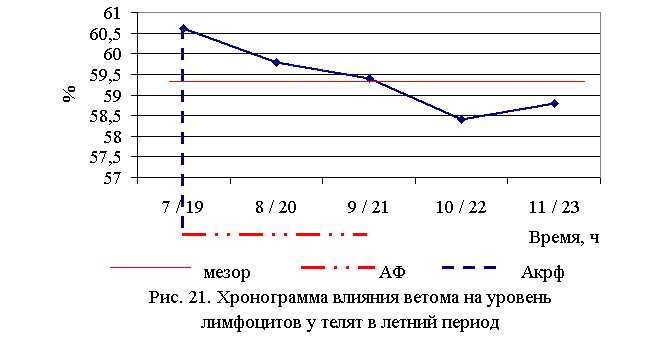
Таблица 16. Лейкоцитарный профиль телят после выздоровления в летний период, %
| Показатель |
Опытные группы |
Контроль |
||||
|
1-я |
2-я |
3-я |
4-я |
5-я |
||
|
Э |
0,6±0,24 Cv=31,29 |
0,4±0,24 Cv=136,93 |
0 |
0,8±0,37 Cv=37,27 |
1,2±0,58 Cv=108,65 |
1,6±0,4 Cv=89,07 |
|
Б |
0 |
0 |
0 |
0 |
0 |
0 |
|
Ю |
1,2±0,2 Cv=20,33 |
1,6±0,24 Cv=34,23 |
1,2±0,58 Cv=108,65 |
1,2±0,58 Cv=108,65 |
1,2±0,49 Cv=91,29 |
0,6±0,4 Cv=136,93 |
|
П |
5,4±0,24 Cv=7,40 |
6,2±0,2 Cv=7,21 |
6,4±0,51 Cv=17,82 |
6,4±0,24 Cv=8,56 |
6,6±0,4 Cv=13,55 |
7,6±0,4 Cv=13,55 |
|
С |
28,4±1,08 Cv=7,67 |
29,2±0,37 Cv=2,68 |
30,4±0,81 Cv=5,98 |
30,0±0,55 Cv=3,83 |
28,4±0,6 Cv=4,02 |
31,2±0,66 Cv=4,61 |
|
Л |
60,6±1,4* Cv=5,75 |
59,8±0,37** Cv=1,45 |
59,4±1,61 Cv=6,11 |
58,4±1,63 Cv=6,47 |
58,8±0,73 Cv=3,05 |
56,8±1,24 Cv=4,97 |
|
М |
3,8±0,37** Cv=22,02 |
2,8±0,49 Cv=39,12 |
2,6±0,4 Cv=34,4 |
3,2±0,8 Cv=55,9 |
3,8±0,49* Cv=28,83 |
2,2±0,58 Cv=40,75 |
При применении ветома 3 уровень моноцитов у телят в летний период также изменялся. В летний период процентное содержание моноцитов у телят 1–5-й опытных групп было выше после выздоровления по сравнению с аналогами из контроля на 72,7 (P<0,05); 27,3; 18,2; 45,5; 72,7 (P<0,1) % соответственно. По сравнению с исходными данными уровень моноцитов снизился у телят 2–5-й опытных и контрольной групп на 12,5; 13,3; 11,1; 5,0; 47,6 %, тогда как у животных 1-й опытной группы – увеличился на 46,2 %. Необходимо отметить, что все изменения в уровне моноцитов происходили в пределах физиологической нормы (табл. 15-16).
Под влиянием препарата происходили изменения в соотношении нейтрофилов у телят.
До лечения в летний период среди видового состава нейтрофилов преобладали сегментоядерные клетки, их количество соответствовало верхней границе нормы, как в опытных, так и в контрольной группе.
После выздоровления по сравнению с аналогами из контроля у телят 1–5-й групп уровень сегментоядерных нейтрофилов был ниже на 9,0; 6,4; 2,6; 3,8; 9,0 %, а количество палочкоядерных нейтрофилов – на 28,9; 18,4; 15,8; 15,8; 13,2 %. Процентное содержание юных форм у телят 1, 3, 4 и 5-й опытных групп превышало аналогов из контроля выше на 25,0 %, а у животных 2-й опытной группы – находилось на уровне контрольной группы.
По сравнению с исходными данными у телят 1–5-й опытных групп уровень сегментоядерных нейтрофилов уменьшился на 12,3; 9,9; 2,6; 8,5; 9,6% соответственно, тогда как у аналогов из контроля – увеличился на 0,6%. Количество палочкоядерных форм снизилось у телят 1, 2 и 3-й опытных групп на 12,9; 13,9 и 3,0 %, а у животных 4-й, 5-й опытных и контрольной групп – увеличилось на 10,3; 6,5 и 2,7 % соответственно. Содержание юных нейтрофилов увеличилось у телят 1–5-й опытных групп на 50,0; 33,3; 50,0; 50,0; 20,0 % соответственно, тогда как у аналогов из контроля – не изменилось по сравнению с исходными данными (табл. 15-16).
При проведении опыта в летний период отмечены изменения в уровне эозинофилов у телят.
Количество эозинофилов у телят 1, 2, 4 и 5-й опытных групп было выше по сравнению с аналогами из контроля на 62,3; 75,0; 50,0; 25,0 % соответственно, тогда как у телят 3-й опытной группы при анализе лейкограммы эозинофилов обнаружено не было. По сравнению с исходными данными уровень эозинофилов увеличился у телят 1, 4, 5-й опытных и контрольной групп в 1,5; 2,0; 2,0 и 1,3 раз, а у животных 2-й опытной группы – уменьшился в 0,5 раза. Анализ хронобиологических особенностей влияния ветома 3 на уровень эозинофилов не позволяет сделать однозначных выводов (табл. 15-16).
При проведении опыта в осенний период процентное соотношение лейкоцитарных клеток у телят также изменялось.
Процентное содержание лимфоцитов после выздоровления в осенний период у телят 1–5-й опытных групп было выше по сравнению с аналогичным показателем животных из контроля на 2,1; 2,4; 2,3; 1,7; 1,0 % соответственно. По сравнению с исходными данными содержание лимфоцитов увеличилось у телят 1–5-й опытных и контрольной групп на 1,7; 5,8; 4,4; 2,3; 1,4 и 0,7 % соответственно (табл. 17-18).
В активной фазе влияния ветома 3 на уровень лимфоцитов в осенний период находились показатели 1, 2 и 3-й опытных групп, которые превышали мезор (58,5±0,14%) на 0,2; 0,5 и 0,3 %, в пассивной фазе – показатели 4-й и 5-й опытных групп, которые были ниже мезора на 0,2 и 0,9 % соответственно. Продолжительность активной фазы в 2 раза превышала пассивную. Абсолютная амплитуда показателей уровня лимфоцитов составила 0,8 %, относительная амплитуда – 1,01 ед. Коэффициент синхронизации влияния ветома 3 на содержание лимфоцитов у телят в осенний период составил 0,34 ед./ч. Акрофаза влияния ветома 3 на уровень лимфоцитов у телят приходилась на 8 и 20 ч, что соответствует показателям телят 2-й опытной группы (рис. 22).
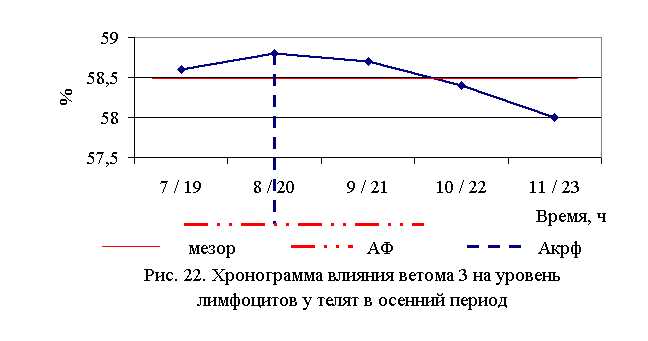
Таблица 17. Лейкоцитарный профиль телят до лечения в осенний период, %
| Показатель |
Опытные группы |
Контроль |
||||
|
1-я |
2-я |
3-я |
4-я |
5-я |
||
|
Э |
0,4±0,24 Cv=136,93 |
0,6±0,24 Cv=91,29 |
0,6±0,24 Cv=91,29 |
0,4±0,24 Cv=136,93 |
1,2±0,97 Cv=180,66 |
0,4±0,24 Cv=136,93 |
|
Б |
0 |
0 |
0 |
0 |
0 |
0 |
|
Ю |
1,8±0,37 Cv=104,58 |
1,8±0,37 Cv=104,58 |
2,0±0,45 Cv=100,0 |
1,0±0,32 Cv=70,71 |
1,0±0,45 Cv=100,0 |
0,8±0,58 Cv=162,98 |
|
П |
6,0±0,71 Cv=26,35 |
7,0±0,32 Cv=11,79 |
6,4±0,75 Cv=26,15 |
6,5±0,8 Cv=26,31 |
5,7±1,12 Cv=46,48 |
5,7±0,55 Cv=20,41 |
|
С |
31,6±1,07 Cv=7,51 |
32,0±0,71 Cv=4,94 |
31,8±0,93 Cv=6,56 |
32,1±0,97 Cv=6,82 |
31,4±5,23 Cv=44,3 |
32,2±1,02 Cv=7,08 |
|
Л |
57,6±1,75 Cv=6,79 |
55,6±1,03 Cv=4,14 |
56,2±1,79 Cv=7,14 |
57,1±1,71 Cv=6,75 |
57,2±5,24 Cv=22,44 |
57,0±0,89 Cv=3,51 |
|
М |
2,6±0,6 Cv=37,27 |
3,0±0,32 Cv=14,14 |
3,0±0,68 Cv=34,47 |
2,9±0,75 Cv=49,22 |
3,5±4,01 Cv=64,11 |
3,9±0,68 Cv=42,13 |
Таблица 18. Лейкоцитарный профиль телят после выздоровления в осенний период, %
| Показатель |
Опытные группы |
Контроль |
||||
|
1-я |
2-я |
3-я |
4-я |
5-я |
||
|
Э |
0,4±0,24 Cv=136,93 |
0,6±0,24 Cv=91,29 |
0,6±0,4 Cv=149,07 |
0 |
0,8±0,37 Cv=104,58 |
0 |
|
Б |
0 |
0 |
0 |
0 |
0 |
0 |
|
Ю |
2,6±0,51* Cv=24,79 |
2,0±0,37 Cv=26,15 |
2,1±0,49 Cv=34,23 |
1,8±0,58 Cv=46,57 |
1,2±0,58 Cv=59,27 |
1,2±0,4 Cv=34,4 |
|
П |
7,4±0,51* Cv=15,41 |
6,0±0,32 Cv=11,79 |
6,5±0,49 Cv=17,67 |
6,2±0,84 Cv=37,42 |
6,6±0,97 Cv=45,17 |
5,8±0,49 Cv=22,82 |
|
С |
28,0±0,55*** Cv=4,08 |
29,2±0,58** Cv=4,18 |
29,0±0,84** Cv=6,03 |
30,6±0,75 Cv=5,3 |
28,8±5,45 Cv=43,86 |
32,7±0,98 Cv=6,98 |
|
Л |
58,6±1,03 Cv=4,22 |
58,8±1,44 Cv=5,77 |
58,7±0,75 Cv=3,01 |
58,4±1,21 Cv=4,71 |
58,0±5,51 Cv=24,66 |
57,4±0,93 Cv=3,61 |
|
М |
3,0±0,55 Cv=40,82 |
3,4±0,8 Cv=55,9 |
3,1±0,4 Cv=26,31 |
3,0±0,84 Cv=62,36 |
4,6±3,1 Cv=82,54 |
2,9±0,37 Cv=26,15 |
В осенний период процентное содержание моноцитов у телят 1–5-й опытных групп было выше после выздоровления по сравнению с аналогами из контроля на 3,4; 17,2; 6,9; 3,4; 58,6 %, а по сравнению с исходными данными – на 15,4; 13,3; 3,3; 3,4; 31,4 % соответственно. Уровень моноцитов у телят контрольной группы снизился по сравнению с исходными данными на 25,6 %. Необходимо отметить, что все изменения в уровне моноцитов происходили в пределах физиологической нормы (табл. 17-18).
Под влиянием препарата происходили изменения в соотношении нейтрофилов у телят.
До лечения препаратом в осенний период среди видового состава нейтрофилов преобладали сегментоядерные клетки, их количество соответствовало верхней границе нормы, как в опытных, так и в контрольной группе.
После выздоровления по сравнению с аналогами из контроля у телят 1–5-й групп уровень сегментоядерных нейтрофилов был ниже на 14,4 (P<0,01); 10,7 (P<0,05); 11,3 (P<0,05); 6,4; 11,9 %, количество палочкоядерных нейтрофилов – выше на 27,6 (P<0,1); 3,4; 12,1; 6,9; 13,8 % соответственно. Содержание юных форм нейтрофилов у телят 1-й и 4-й опытных групп было выше аналогов из контроля на 18,2 (P<0,1) и 18,2 %, у животных 2-й и 3-й опытных групп – ниже на 9,1 и 4,5 % соответственно, а у аналогов из 5-й опытной группы – на уровне телят контрольной группы.
По сравнению с исходными данными у телят 1–5-й опытных групп уровень сегментоядерных нейтрофилов уменьшился на 11,4; 8,8; 8,8; 4,7; 8,3% соответственно, тогда как у аналогов из контроля – увеличился на 1,6%. Количество палочкоядерных форм у телят 1, 3, 5-й опытных и контрольной групп увеличилось на 23,3; 1,6; 15,8 и 1,8 %, а у животных 2-й и 5-й опытных групп – снизилось на 14,3 и 4,6 % соответственно. Содержание юных нейтрофилов увеличилось у телят 1–5-й опытных и контрольной групп на 44,4; 11,1; 5,0; 80,0; 20,0 и 50,0 % соответственно (табл. 17-18).
Изменения в уровне нейтрофилов, как летом, так и осенью происходили в пределах физиологической нормы и носили, скорее всего, перераспределительный характер, связанный с процессом выздоровления животных, поэтому анализ хронобиологических особенностей влияния ветома 3 на уровень нейтрофилов не позволяет сделать однозначных выводов.
Уровень эозинофилов у телят, при проведении опыта в осенний период, по сравнению с началом опыта изменился незначительно (табл. 17-18).
Таким образом, полученные результаты свидетельствуют о позитивном влиянии ветома 3 на эритро- и лейкопоэз, в т. ч. лимфоцитопоэз у телят опытных групп, как в летний, так и в осенний периоды исследований. Ветом 3 повышает уровень моноцитов и юных форм нейтрофилов в пределах физиологической нормы, что может свидетельствовать об усилении фагоцитарной функции у телят и разрешении воспалительного процесса. Установлена зависимость влияния ветома 3 на морфологические показатели крови телят от времени применения препарата и сезона. Оптимальное влияние ветома 3 на эритро- и лейкопоэз отмечено при применении препарата в 7 и 19 ч – летом и в 8 и 20 ч – осенью.
3.3. Хронобиологические особенности действия ветома 3 на биохимические показатели крови у телят при гастроэнтерите
При проведении исследований отмечено влияние препарата на белковый обмен у телят опытных групп.
Уровень общего белка его фракций до применения препарата находились в пределах физиологической нормы (табл. 19).
Содержание общего белка у телят 1–5-й опытных групп после выздоровления увеличилось по сравнению с аналогами из контроля на 4,14 (P<0,1); 3,44; 4,14 (P<0,1); 2,85; 2,33 % в летний период и на 1,84; 3,89 (P<0,1); 2,02; 0,71; 1,77 % в осенний период соответственно. По сравнению с исходными данными уровень общего белка у телят 1–5-й опытных и контрольной групп увеличился на 5,12; 4,45; 4,12; 3,16; 3,17 и 0,83 % в летний период и на 3,18; 4,83; 2,38; 1,25; 2,61 и 0,53 % в осенний период соответственно (табл. 19).
Таблица 19. Динамика уровня общего белка у телят, г/л
| Группа |
Лето |
Осень |
||
|
До лечения |
После выздоровления |
До лечения |
После выздоровления |
|
|
1-я опытная |
56,67±0,664 Cv=2,03 |
59,57±0,318* Cv=0,92 |
55,7±0,517 Cv=1,61 |
57,47±0,384 Cv=1,16 |
|
2-я опытная |
56,83±0,176 Cv=0,54 |
59,17±0,296 Cv=0,87 |
55,93±0,145 Cv=0,45 |
58,63±0,317* Cv=1,10 |
|
3-я опытная |
57,03±0,467 Cv=1,42 |
59,57±0,406* Cv=1,18 |
56,23±0,769 Cv=2,37 |
57,57±0,260 Cv=0,78 |
|
4-я опытная |
57,03±0,649 Cv=1,97 |
58,83±0,664 Cv=1,96 |
56,13±0,865 Cv=2,67 |
56,83±0,736 Cv=2,24 |
|
5-я опытная |
56,73±0,426 Cv=1,30 |
58,53±0,371 Cv=1,10 |
55,97±1,241 Cv=3,84 |
57,43±1,027 Cv=3,10 |
|
Контроль |
56,73±0,524 Cv=1,60 |
57,20±0,987 Cv=2,99 |
56,13±0,845 Cv=2,61 |
56,43±0,809 Cv=2,48 |
В активной фазе влияния ветома 3 на уровень общего белка в летний период находились показатели 1, 2 и 3-й опытных групп, которые превышали мезор ((59,13±0,2) г/л) на 0,7; 0,1 и 0,7 %, в пассивной фазе – показатели 4-й и 5-й опытных групп, которые были ниже мезора на 0,5 и 1,0 % соответственно. Продолжительность активной фазы превышала пассивную на 66,7 %. Абсолютная амплитуда показателей уровня общего белка составила 1,04 г/л, относительная амплитуда – 1,02 ед., коэффициент синхронизации – 0,51ед./ч. Акрофаза влияния ветома 3 на уровень общего белка у телят в летний период приходилась на 7 и 19, и 9 и 21 ч, что соответствует показателям телят 1-й и 3-й опытных групп (рис. 23).
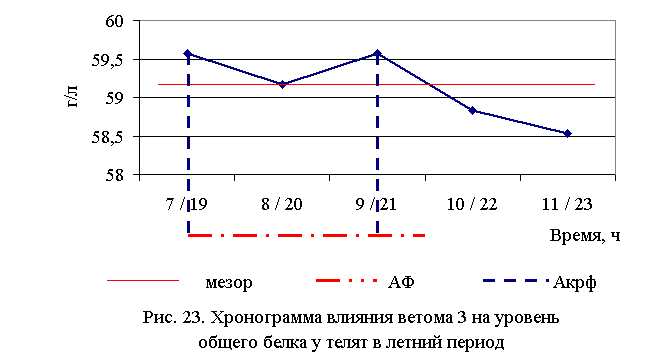
В активной фазе влияния ветома 3 на уровень общего белка в осенний период находились показатели 2-й опытной группы, которые превышали мезор ((57,59±0,29) г/л) на 1,8 %, в пассивной фазе – показатели 1, 3, 4 и 5-й опытных групп, которые были ниже мезора на 0,2; 0,03; 1,3 и 0,3 % соответственно. Продолжительность активной фазы была меньше пассивной на 28,6 %. Абсолютная амплитуда показателей уровня общего белка составила 1,8 г/л, относительная амплитуда – 1,03 ед., коэффициент синхронизации – 0,52 ед./ч. Акрофаза влияния ветома 3 на уровень общего белка у телят в осенний период приходилась на 8 и 20 ч, что соответствует показателям телят 2-й опытной группы (рис. 24). Анализ содержания белковых фракций свидетельствует о повышении процентного содержания уровня альбуминов у телят опытных групп (табл. 20-23).
Содержание альбуминов у телят 1–5-й опытных групп после выздоровления было выше аналогичного показателя животных из контроля на 7,5; 5,3; 6,5; 5,4; 2,6 % в летний, и на 5,3; 5,3; 3,5; 3,8; 6,2 % в осенний периоды соответственно. По сравнению с исходными данными уровень альбуминов увеличился у телят 1–5-й опытных и контрольной групп в летний период – на 11,33; 10,84; 10,22; 10,06; 1,85 и 2,56 %, осенью – на 7,1; 7,3; 6,3; 6,1; 9,7 и 5,1 % соответственно.
Уровень α-глобулинов у телят 1–5-й опытных групп после выздоровления был ниже по сравнению с показателями контрольной группы в летний период – на 31,0; 25,6; 28,7; 34,7; 19,3 %, осенью – на 35,6; 30,2; 26,3; 15,3; 18,8 % соответственно. По сравнению с исходными данными уровень α-глобулинов у телят 1–5-й опытных и контрольной групп снизился на 36,1; 36,7; 26,2; 39,0; 7,4 и 0,8 % – в летний и на 42,8; 33,8; 31,8; 17,5; 42,3 и 18,1 % – в осенний периоды соответственно (табл. 20-23).
Содержание β-глобулинов в крови телят 1–5-й опытных групп после выздоровления в летний период было ниже по сравнению с показателями контрольной группы на 11,9; 4,8; 5,4; 3,0; 4,2 % соответственно. При проведении исследований осенью уровень β-глобулинов у телят 1, 2 и 5-й опытных групп превышал показатели контрольной группы на 1,7, 1,2 и 4,3%, а у телят 3 и 4-й опытных групп – был ниже на 1,4 и 4,3% соответственно. По сравнению с исходными данными содержание β-глобулинов снизилось у телят 1–5-й опытных и контрольной групп на 17,4; 8,8; 17,5; 6,9; 10,9 и 8,6 % - в летний, и на 2,2; 6,9; 10,0; 16,5; 7,0 и 7,6 % - в осенний периоды соответственно (табл. 20-23).
Таблица 20. Фракции белка у телят до лечения в летний период, %
| Фракции белка |
Опытные |
контроль |
||||
|
1-я группа |
2-я группа |
3-я группа |
4-я группа |
5-я группа |
||
|
Альбу-мины |
51,83±0,79 Cv=2,63 |
51,0±0,96 Cv=3,28 |
51,87±0,92 Cv=3,07 |
51,4±1,93 Cv=6,50 |
54,07±0,27 Cv=0,87 |
52,33±1,12 Cv=3,71 |
|
α – гло-булины |
18,47±1,83 Cv=17,13 |
20,10±0,76 Cv=6,52 |
16,53±0,57 Cv=5,97 |
18,30±2,40 Cv=22,72 |
14,90±1,64 Cv=19,03 |
17,23±1,03 Cv=10,32 |
|
β – гло-булины |
17,67±0,85 Cv=8,34 |
17,3±0,12 Cv=1,16 |
19,0±1,07 Cv=9,75 |
17,27±0,54 Cv=5,38 |
17,83±1,54 Cv=14,97 |
18,13±0,38 Cv=3,59 |
|
γ – гло-булины |
12,03±0,35 Cv=5,01 |
11,27±0,65 Cv=9,98 |
12,6±0,53 Cv=7,27 |
13,03±0,84 Cv=11,10 |
13,2±0,53 Cv=6,94 |
12,30±1,00 Cv=14,11 |
Таблица 21. Фракции белка у телят после выздоровления в летний период, %
| Фракции белка |
Опытные |
контроль |
||||
|
1-я группа |
2-я группа |
3-я группа |
4-я группа |
5-я группа |
||
|
Альбу-мины |
57,70±1,12 Cv=3,35 |
56,53±0,67 Cv=2,05 |
57,17±0,84 Cv=2,55 |
56,57±0,71 Cv=2,18 |
55,07±0,72 Cv=2,27 |
53,67±0,78 Cv=2,52 |
|
α – гло-булины |
11,80±0,81 Cv=11,86 |
12,73±0,30 Cv=4,03 |
12,20±0,85 Cv=12,13 |
11,17±1,65 Cv=25,56 |
13,80±2,20 Cv=27,65 |
17,10±0,61 Cv=6,19 |
|
β – гло-булины |
14,6±0,40 Cv=4,79 |
15,77±0,64 Cv=6,99 |
15,67±0,52 Cv=5,79 |
16,07±1,26 Cv=13,53 |
15,87±0,35 Cv=3,80 |
16,57±0,32 Cv=3,32 |
|
γ – гло-булины |
15,9±0,47 Cv=5,15 |
14,97±0,74 Cv=8,59 |
14,97±0,38 Cv=4,45 |
16,20±0,44 Cv=4,66 |
15,27±1,82 Cv=20,65 |
12,67±0,20 Cv=2,77 |
Уровень γ-глобулинов у телят 1–5-й опытных групп после выздоровления превышал аналогичный показатель животных контрольной группы в летний период – на 25,5; 18,2; 18,2; 27,9; 20,5 %, осенью – на 16,5; 11,2; 22,2; 10,7; 13,9 % соответственно. По сравнению с исходными данными содержание γ-глобулинов увеличилось у телят 1–5-й опытных и контрольной групп на 32,2; 32,8; 18,8; 24,3; 15,7 и 3,0 % – в летний и на 32,8; 22,6; 35,4; 20,1; 9,9 и 18,2 % – в осенний периоды соответственно (табл. 20-23).
Таблица 22. Фракции белка у телят до лечения в осенний период, %
| Фракции Белка |
Опытные |
Контроль |
||||
|
1-я группа |
2-я группа |
3-я группа |
4-я группа |
5-я группа |
||
|
Альбу-мины |
53,83±0,96 Cv=3,09 |
53,77±0,27 Cv=0,88 |
53,3±1,27 Cv=4,11 |
53,57±0,38 Cv=1,21 |
53,03±0,63 Cv=2,07 |
52,1±0,64 Cv=2,11 |
|
α – гло-булины |
17,43±0,81 Cv=8,04 |
16,33±0,55 Cv=5,79 |
16,73±1,28 Cv=13,26 |
15,87±0,15 Cv=1,59 |
16,4±1,12 Cv=11,78 |
18,9±0,5 Cv=4,58 |
|
β – гло-булины |
16,73±0,84 Cv=8,67 |
17,5±0,46 Cv=4,54 |
17,63±0,45 Cv=4,40 |
17,97±1,02 Cv=9,80 |
18,07±0,27 Cv=2,62 |
17,43±0,24 Cv=2,39 |
|
γ – гло-булины |
12,0±0,99 Cv=14,22 |
12,4±0,25 Cv=3,52 |
12,33±0,81 Cv=11,39 |
12,6±0,66 Cv=9,01 |
14,17±2,07 Cv=25,28 |
11,57±0,55 Cv=8,22 |
Таблица 23. Фракции белка у телят после выздоровления в осенний период, %
| Фракции белка |
Опытные |
Контроль |
||||
|
1-я группа |
2-я группа |
3-я группа |
4-я группа |
5-я группа |
||
|
Альбу-мины |
57,67±0,22 Cv=0,66 |
57,7±0,85 Cv=2,56 |
56,67±0,47 Cv=1,44 |
56,83±0,82 Cv=2,49 |
58,17±0,32 Cv=0,95 |
54,77±1,62 Cv=5,11 |
|
α – гло-булины |
9,97±0,35 Cv=6,05 |
10,8±1,12 Cv=18,03 |
11,4±0,61 Cv=9,28 |
13,1±1,19 Cv=15,77 |
9,47±1,19 Cv=21,71 |
15,47±1,89 Cv=21,11 |
|
β – гло-булины |
16,37±0,54 Cv=5,68 |
16,3±0,31 Cv=3,25 |
15,87±0,54 Cv=5,86 |
15,0±0,25 Cv=2,91 |
16,8±0,23 Cv=2,38 |
16,1±1,32 Cv=14,23 |
|
γ – гло-булины |
15,93±0,24 Cv=2,61 |
15,2±0,55 Cv=6,24 |
16,7±0,67 Cv=7,21 |
15,13±0,91 Cv=10,46 |
15,57±0,81 Cv=9,02 |
13,67±1,50 Cv=19,04 |
При проведении опыта отмечены также изменения в уровне холестерина в крови телят опытных групп. Содержание холестерина у телят 1–5-й опытных групп по сравнению с аналогами из контроля было ниже в летний период – на 5,2 (P<0,001); 3,1 (P<0,01); 2,8 (P<0,05); 0,7; 2,1 (P<0,1) %, осенью – на 1,4 (P<0,1); 1,5; 1,0; 1,0; 1,0 % соответственно. По сравнению с исходными данными в летний период, уровень холестерина снизился у телят 1, 2, 3 и 5-й опытных групп на 4,3; 1,1; 2,0; 1,5 %, а у животных 4-й опытной и контрольной групп – увеличился на 0,5 и 0,4 % соответственно. В осенний период данный показатель снизился у телят 1–5-й опытных и контрольной групп на 1,0; 1,6; 0,9; 1,2; 0,7 и 0,3 % соответственно (табл. 24).
Таблица 24. Динамика уровня холестерина в крови подопытных телят, ммоль/л
| Группа |
Лето |
Осень |
||
|
До лечения |
После выздоровления |
До лечения |
После выздоровления |
|
|
1-я опытная |
4,642±0,037 Cv=1,78 |
4,444±0,021# Cv=1,07 |
4,649±0,027 Cv=1,28 |
4,601±0,011* Cv=0,52 |
|
2-я опытная |
4,596±0,069 Cv=3,37 |
4,544±0,026*** Cv=1,28 |
4,674±0,012 Cv=0,58 |
4,598±0,025 Cv=1,2 |
|
3-я опытная |
4,648±0,057 Cv=2,73 |
4,557±0,038** Cv=1,86 |
4,665±0,010 Cv=0,46 |
4,621±0,012 Cv=0,56 |
|
4-я опытная |
4,628±0,026 Cv=1,27 |
4,652±0,035 Cv=1,69 |
4,673±0,024 Cv=1,15 |
4,619±0,017 Cv=0,82 |
|
5-я опытная |
4,66±0,019 Cv=0,93 |
4,589±0,038* Cv=1,85 |
4,652±0,014 Cv=0,69 |
4,621±0,017 Cv=0,83 |
|
контроль |
4,654±0,012 Cv=0,57 |
4,635±0,028 Cv=1,34 |
4,655±0,011 Cv=0,52 |
4,633±0,014 Cv=0,65 |
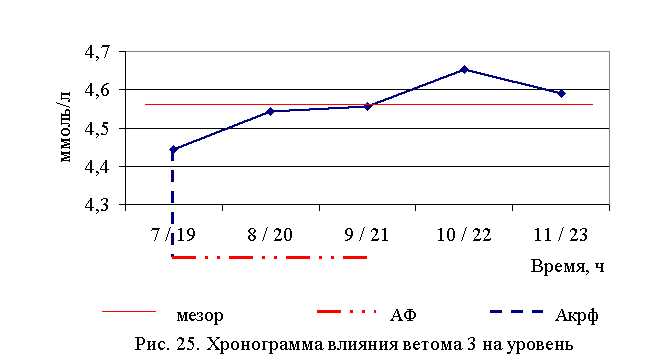
В активной фазе (в данном случае – ниже мезора, т. к. отмечена тенденция к понижению уровня холестерина у телят всех групп) влияния ветома 3 на уровень холестерина в летний период находились показатели 1, 2 и 3-й опытных групп, которые были ниже мезора ((4,567±0,034) ммоль/л) на 2,7; 0,5 и 0,2 %, в пассивной фазе – показатели 4-й и 5-й опытных групп, которые были выше мезора на 1,9 и 0,5 % соответственно. Продолжительность активной фазы превышала пассивную на 18,2 %. Абсолютная амплитуда показателей уровня холестерина составила 0,21 ммоль/л, относительная амплитуда – 1,05 ед., коэффициент синхронизации – 0,35 ед./ч. Акрофаза влияния ветома 3 на уровень холестерина у телят в летний период приходилась на 7 и 19 ч, что соответствует показателям телят 1-й опытной группы (рис. 25).
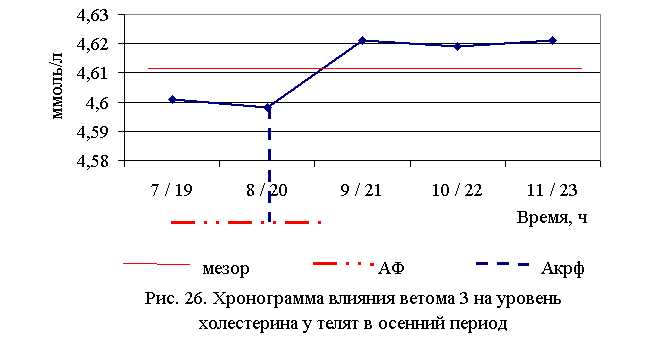
В активной фазе влияния ветома 3 на уровень холестерина в осенний период находились показатели 1-й и 2-й опытных групп, которые были ниже мезора ((4,612±0,005) ммоль/л) на 0,2 и 0,3 %, в пассивной фазе – показатели 3, 4 и 5-й опытных групп, которые были выше мезора на 0,2 % соответственно. Продолжительность активной фазы была меньше пассивной на 54,5 %. Абсолютная амплитуда показателей уровня холестерина составила 0,02 ммоль/л, относительная амплитуда – 1,01 ед., коэффициент синхронизации – 1,01 ед./ч. Акрофаза влияния ветома 3 на уровень холестерина у телят в осенний период приходилась на 8 и 20 ч, что соответствует показателям телят 2-й опытной группы (рис. 26). Таким образом, ветом 3 обладает выраженным влиянием на показатели белкового обмена, повышая уровень общего белка, альбуминов и g-глобулинов в пределах физиологических норм. При применении препарата также отмечено понижение уровня холестерина. Снижение показателей a- и b-глобулинов, скорее всего, связано с разрешением воспалительного процесса. Установлена зависимость влияния ветома 3 на показатели белкового обмена от временного фактора. Оптимальное повышение уровня общего белка и снижение уровня холестерина отмечено при применении препарата в 7 и 19 ч – в летний период, и в 8 и 20 ч – осенью.
При проведении исследований отмечены изменения в уровне небелковых азотистых компонентов крови телят опытных групп. Показатели уровня мочевины и креатинина в крови животных всех групп находились в пределах физиологической нормы.
Под влиянием ветома 3 уровень мочевины в крови телят опытных групп достоверно снижался (табл. 25).
Содержание мочевины в крови телят 1–5-й опытных групп по сравнению с аналогами из контроля было ниже в летний период – на 13,4 (P<0,001); 9,1 (P<0,1); 1,2; 5,2 (P<0,1); 2,8 %, осенью – на 2,8 (P<0,01); 4,4 (P<0,05); 3,8 (P<0,05); 1,8 (P<0,1); 1,8 % соответственно. По сравнению с исходными данными в летний период уровень мочевины снизился у телят 1–5-й опытных групп на 10,0; 9,4; 2,1; 5,7; 3,8 % соответственно, а у аналогов из контроля – увеличился на 0,8 %. В осенний период данный показатель снизился у телят 1–5-й опытных и контрольной групп на 6,0; 6,1; 4,9; 6,3; 3,6 и 1,8 % соответственно (табл. 25).
Таблица 25. Динамика уровня мочевины в крови подопытных телят, ммоль/л
| Группа |
Лето |
Осень |
||
|
До лечения |
После выздоровления |
До лечения |
После выздоровления |
|
|
1-я опытная |
12,841±0,188 Cv=3,27 |
11,551±0,267# Cv=5,17 |
13,594±0,237 Cv=3,89 |
12,771±0,012*** Cv=2,01 |
|
2-я опытная |
13,381±0,399 Cv=6,66 |
12,118±0,547* Cv=10,09 |
13,363±0,148 Cv=2,48 |
12,553±0,207** Cv=3,69 |
|
3-я опытная |
13,451±0,271 Cv=4,51 |
13,169±0,182 Cv=3,09 |
13,274±0,234 Cv=3,94 |
12,630±0,135** Cv=2,38 |
|
4-я опытная |
13,401±0,219 Cv=3,65 |
12,637±0,291* Cv=5,15 |
13,767±0,136 Cv=2,22 |
12,902±0,074* Cv=1,29 |
|
5-я опытная |
13,468±0,135 Cv=2,24 |
12,962±0,211 Cv=3,64 |
13,376±0,152 Cv=2,55 |
12,896±0,186 Cv=3,22 |
|
контроль |
13,711±0,254 Cv=4,14 |
12,944±0,255 Cv=4,41 |
13,202±0,285 Cv=4,82 |
12,754±0,229 Cv=4,01 |
В активной фазе (в данном случае – ниже мезора, т.к. во время исследований отмечено снижение уровня мочевины во всех группах) влияния ветома 3 на уровень мочевины в летний период находились показатели 1-й и 2-й опытных групп, которые были ниже мезора ((12,487±0,29) ммоль/л) на 7,5 и 3,0 %, в пассивной фазе – показатели 3, 4 и 5-й опытных групп, которые были выше мезора на 5,5; 1,2 и 3,8 % соответственно. Продолжительность активной фазы была короче пассивной на 54,5 %. Абсолютная амплитуда показателей уровня мочевины составила 1,6 ммоль/л, относительная амплитуда – 1,14 ед., коэффициент синхронизации – 0,57 ед./ч. Акрофаза влияния ветома 3 на уровень мочевины у телят в летний период приходилась на 7 и 19 ч, что соответствует показателям телят 1-й опытной группы (рис. 27). В активной фазе влияния ветома 3 на уровень мочевины в осенний период находились показатели 2-й и 3-й опытных групп, которые были ниже мезора ((12,76±0,29) ммоль/л) на 1,5 и 0,9 %, в пассивной фазе – показатели 1, 4 и 5-й опытных групп, которые были выше мезора на 0,2; 1,2 и 1,1 % соответственно. Продолжительность активной фазы превышала пассивную в 2 раза. Абсолютная амплитуда показателей уровня мочевины составила 0,35 ммоль/л, относительная амплитуда – 1,03 ед., коэффициент синхронизации – 0,52 ед./ч. Акрофаза влияния ветома 3 на уровень мочевины у телят в осенний период приходилась на 8 и 20 ч, что соответствует показателям телят 2-й опытной группы (рис. 28).
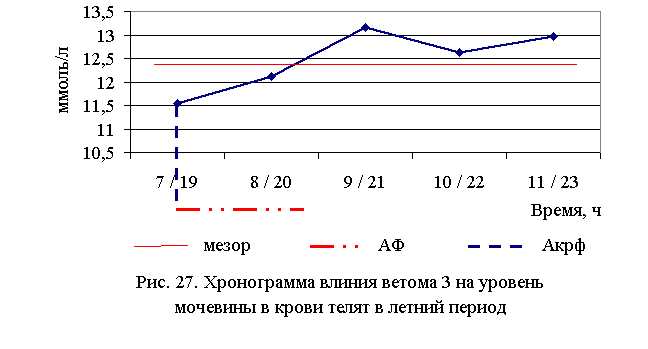
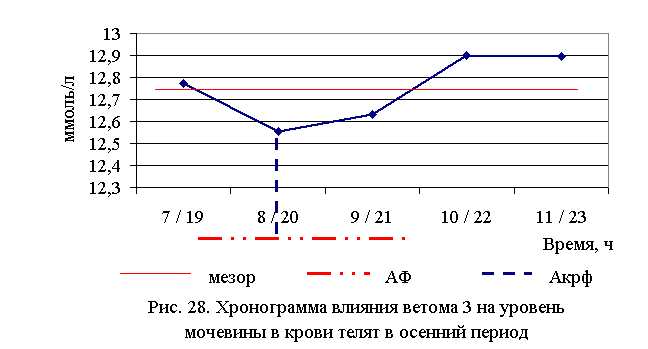
При проведении исследований уровень креатинина в крови телят опытных групп также понижался (табл. 26).
Содержание креатинина после выздоровления в крови животных 1–5-й опытных было ниже по сравнению с аналогами из контроля в летний период – на 27,1; 20,3; 20,9; 14,9; 22,3 %, осенью – на 16,7; 25,6; 21,1; 18,1; 6,7 % соответственно. По сравнению с исходными данными уровень креатинина в крови телят 1–5-й опытных снизился на 16,3; 3,8; 10,0; 8,2; 3,1 % – в летний, и на 15,2; 27,2; 14,9; 13,0; 2,6 % – в осенний периоды соответственно. Тогда как у аналогов из контроля количество креатинина в крови увеличилось по сравнению с исходными данными на 7,9 и 0,4 % летом и осенью соответственно (табл. 26).
Таблица 26. Динамика уровня креатинина в крови подопытных телят, мкмоль/л
| Группа |
Лето |
Осень |
||
|
До лечения |
После выздоровления |
До лечения |
После выздоровления |
|
|
1-я опытная |
20,684±0,734 Cv=6,15 |
17,313±1,16 Cv=11,60 |
22,363±0,794 Cv=6,15 |
18,971±0,886 Cv=8,08 |
|
2-я опытная |
19,692±2,204 Cv=19,39 |
18,945±0,922 Cv=8,44 |
23,258±1,84 Cv=13,7 |
16,941±1,255 Cv=12,83 |
|
3-я опытная |
20,88±2,327 Cv=19,3 |
18,799±1,22 Cv=11,24 |
21,093±3,555 Cv=29,19 |
17,957±1,851 Cv=17,86 |
|
4-я опытная |
22,029±3,253 Cv=25,58 |
20,225±1,535 Cv=13,14 |
21,446±0,763 Cv=6,16 |
18,654±1,094 Cv=10,16 |
|
5-я опытная |
19,039±0,95 Cv=8,64 |
18,455±1,035 Cv=9,72 |
21,807±2,769 Cv=21,99 |
21,25±1,81 Cv=14,75 |
|
контроль |
21,684±5,202 Cv=41,55 |
20,268±3,264 Cv=27,9 |
21,631±0,492 Cv=3,94 |
19,843±0,331 Cv=2,89 |
В активной фазе (ниже мезора) влияния ветома 3 на уровень креатинина в летний период находились показатели 1-й и 5-й опытных групп, которые были ниже мезора ((18,747±0,467) мкмоль/л) на 7,6 и 1,6 %, в пассивной фазе – показатели 2, 3 и 4-й опытных групп, которые были выше мезора на 1,1; 0,3 и 7,9 % соответственно. Продолжительность активной фазы была меньше пассивной на 70,3 %. Абсолютная амплитуда показателей уровня креатинина составила 2,9 мкмоль/л, относительная амплитуда – 1,2 ед., коэффициент синхронизации – 0,4 ед./ч. Акрофаза влияния ветома 3 на уровень креатинина у телят в летний период приходилась на 7 и 19 ч, что соответствует показателям телят 1-й опытной группы (рис. 29).
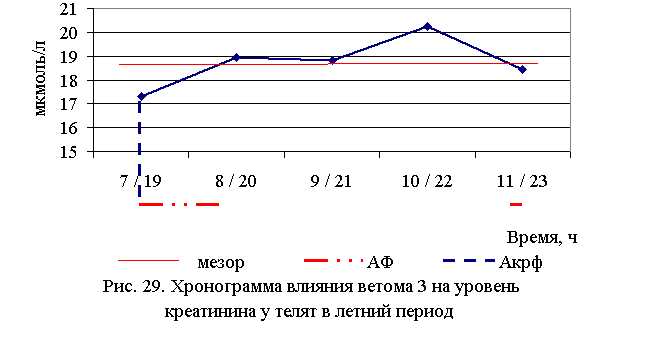
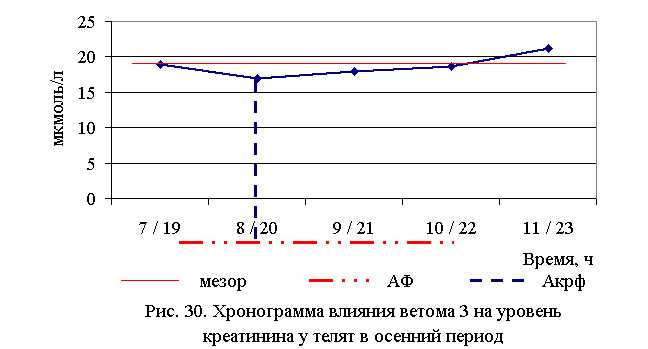
В активной фазе влияния ветома 3 на уровень креатинина в осенний период находились показатели 2, 3 и 4-й опытных групп, которые были ниже мезора ((18,755±0,714) мкмоль/л) на 9,7; 4,3 и 0,5 %, в пассивной фазе – показатели 1-й и 5-й опытных групп, которые были выше мезора на 1,2 и 13,3% соответственно. Продолжительность активной фазы превышала пассивную на 66,7 %. Абсолютная амплитуда показателей уровня креатинина составила 4,3 мкмоль/л, относительная амплитуда – 1,3 ед., коэффициент синхронизации – 0,43 ед./ч. Акрофаза влияния ветома 3 на уровень креатинина у телят в осенний период приходилась на 8 и 20 ч, что соответствует показателям телят 2-й опытной группы (рис. 30). Таким образом, ветом 3 обладает выраженным влиянием на небелковые азотистые компоненты крови, вызывая понижение уровня мочевины и креатинина у телят, что может свидетельствовать об улучшении детоксицирующей функции печени. Оптимальное снижение уровня мочевины и креатинина отмечено при применении препарата в 7 и 19 ч – в летний период, и в 8 и 20 ч – осенью.
При проведении опыта отмечены изменения в уровне желчных пигментов в крови телят. Необходимо отметить, что до лечения уровень билирубина и его фракций превышал границу физиологической нормы во всех группах. При применении ветома 3 наблюдалась тенденция к понижению уровня данных показателей в крови телят опытных групп (табл. 27-32).
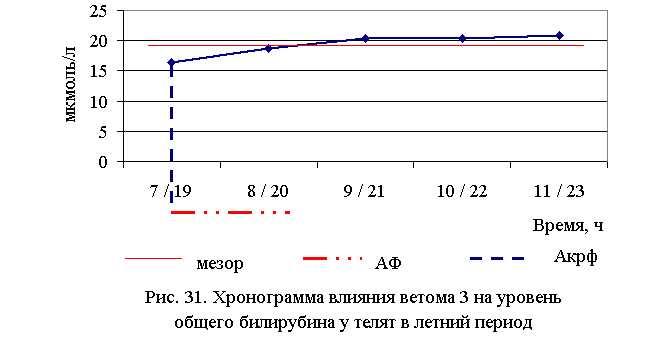
Содержание общего билирубина в крови телят 1–5-й опытных групп после выздоровления был ниже по сравнению с аналогами из контроля в летний период – на 24,4 (P<0,1); 14,3 (P<0,1); 6,3; 6,1; 4,3 %, осенью – на 8,56; 19,57 (P<0,001); 9,70 (P<0,05); 4,96; 4,74 % соответственно. По сравнению с исходными данными уровень общего билирубина снизился у телят 1–5-й опытных и контрольной групп на 29,0; 17,5; 6,6; 5,9; 12,3 и 0,3 % – в летний, и на 10,06; 21,33; 9,54; 5,20; 5,44 и 0,52 % – в осенний периоды соответственно (табл. 27).
Таблица 27. Динамика содержания общего билирубина в крови телят в летний период, мкмоль/л
| Группа |
До лечения |
После выздоровления |
|
1-я опытная |
23,47±0,90 Cv=8,6 |
16,45±1,04** Cv=14,13 |
|
2-я опытная |
22,60±1,08 Cv=10,66 |
18,65±0,67* Cv=8,01 |
|
3-я опытная |
21,84±1,63 Cv=16,32 |
20,40±0,37 Cv=4,08 |
|
4-я опытная |
21,73±2,15 Cv=22,09 |
20,44±0,89 Cv=9,71 |
|
5-я опытная |
23,75±0,48 Cv=4,5 |
20,84±1,2 Cv=12,87 |
|
Контроль |
21,94±0,88 Cv=8,97 |
20,45±1,22 Cv=13,37 |
Таблица 28. Динамика содержания общего билирубина в крови телят в осенний период, мкмоль/л
| Группа |
До лечения |
После выздоровления |
|
1-я опытная |
23,17±0,88 Cv=8,51 |
20,84±1,03 Cv=11,05 |
|
2-я опытная |
23,3±0,84 Cv=8,08 |
18,33±0,55# Cv=6,66 |
|
3-я опытная |
22,75±0,68 Cv=6,68 |
20,58±0,77** Cv=8,38 |
|
4-я опытная |
22,83±0,94 Cv=9,21 |
21,66±0,80 Cv=8,25 |
|
5-я опытная |
22,96±0,66 Cv=6,42 |
21,71±0,57 Cv=5,86 |
|
Контроль |
23,45±0,49 Cv=4,74 |
21,63±0,74 Cv=7,62 |
В активной фазе (ниже мезора) влияния ветома 3 на уровень общего билирубина в летний период находились показатели 1-й и 2-й опытных групп, которые были ниже мезора ((19,356±0,819) мкмоль/л) на 15,0 и 3,6 %, в пассивной фазе – показатели 3, 4 и 5-й опытных групп, которые были выше мезора на 5,4; 5,6 и 7,7 % соответственно. Продолжительность активной фазы была меньше пассивной на 54,5%. Абсолютная амплитуда показателей уровня общего билирубина составила 4,4 мкмоль/л, относительная амплитуда – 1,3 ед., коэффициент синхронизации – 0,33 ед./ч. Акрофаза влияния ветома 3 на уровень общего билирубина у телят в летний период приходилась на 7 и 19 ч, что соответствует показателям телят 1-й опытной группы (рис. 31).
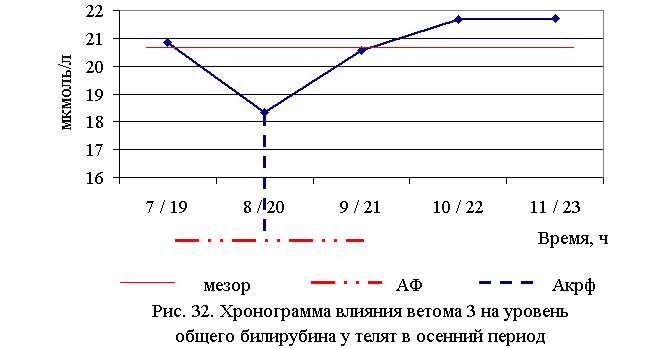
В активной фазе влияния ветома 3 на уровень общего билирубина в осенний период находились показатели 2-й и 3-й опытных групп, которые были ниже мезора ((20,624±0,615) мкмоль/л) на 11,1 и 0,2 %, в пассивной фазе – показатели 1, 4 и 5-й опытных групп, которые были выше мезора на 1,0; 5,0 и 5,3 % соответственно. Продолжительность активной и пассивной фаз были равными. Абсолютная амплитуда показателей уровня общего белка составила 3,4 мкмоль/л, относительная амплитуда – 1,2 ед., коэффициент синхронизации – 0,4 ед./ч. Акрофаза влияния ветома 3 на уровень общего билирубина у телят в осенний период приходилась на 8 и 20 ч, что соответствует показателям телят 2-й опытной группы (рис. 32).
После выздоровления в крови телят 1–5-й опытных групп уровень конъюгированного билирубина был ниже по сравнению с аналогами из контроля в летний период – на 33,3; 21,2; 15,5; 11,2; 15,0 %, осенью – на 15,6; 25,3; 12,5; 7,2; 6,1 % соответственно. По сравнению с исходными данными содержание конъюгированного билирубина в крови телят 1–5-й опытных и контрольной групп снизилось на 25,6; 21,2; 11,3; 8,9; 14,5 и 2,7 % – в летний, и на 20,6; 31,8; 18,2; 16,0; 15,0 и 6,84 % – в осенний периоды соответственно (табл. 29-30). Уровень неконъюгированного билирубина в крови телят 1-й и 2-й опытных групп после выздоровления в летний период был ниже показателей контрольной группы на 11,5 и 4,2 %, а у животных 3, 4 и 5-й опытных групп – выше на 7,3; 1,4 и 11,5 % соответственно. В осенний период у телят 1–5-й опытных групп данный показатель был ниже уровня неконъюгированного билирубина телят из контроля на 1,0; 13,5; 6,8; 2,6; 3,3 % соответственно.
Таблица 29. Динамика уровня конъюгированного билирубина у телят в летний период, мкмоль/л
| Группа |
До лечения |
После выздоровления |
|
1-я опытная |
11,61±1,02 Cv=19,65 |
8,64±0,56# Cv=14,49 |
|
2-я опытная |
12,94±0,75 Cv=12,91 |
10,2±0,64** Cv=14,0 |
|
3-я опытная |
12,33±1,33 Cv=24,17 |
10,94±0,92* Cv=18,8 |
|
4-я опытная |
12,63±1,15 Cv=20,38 |
11,5±0,38* Cv=7,41 |
|
5-я опытная |
12,88±0,67 Cv=11,64 |
11,01±1,32 Cv=26,83 |
|
Контроль |
13,31±0,46 Cv=7,65 |
12,95±0,53 Cv=9,12 |
По сравнению с исходными данными, в летний период содержание неконъюгированного билирубина снизилось у телят 1–5-й опытных групп на 34,1; 12,5; 1,6; 1,8; 9,6 % соответственно, тогда как у аналогов из контроля данный показатель увеличился на 1,1 % (табл. 31-32). В осенний период уровень неконъюгированного билирубина у телят 2-й опытной группы был ниже по сравнению с исходными данными на 8,6 %, тогда как у животных 1, 3, 4, 5-й опытных и контрольной групп данный показатель увеличился на 2,3; 1,0; 9,0; 6,8 и 7,2 % соответственно (табл. 31-32).
Таким образом, ветом 3 оказывает влияние на показатели пигментного обмена, понижая уровень билирубина и его фракций в крови телят опытных групп, что может свидетельствовать об улучшении функции печени. Установлена зависимость влияния пробиотика на уровень билирубина в зависимости от времени применения препарата. Максимальное снижение общего билирубина и его фракций отмечено при применении ветома 3 в 7 и 19 ч – в летний период, и в 8 и 20 ч – осенью.
Таблица 30. Динамика уровня конъюгированного билирубина у телят в осенний период, мкмоль/л
| Группа |
До лечения |
После выздоровления |
|
1-я опытная |
12,46±0,63 Cv=11,32 |
9,89±0,63* Cv=14,3 |
|
2-я опытная |
12,82±0,37 Cv=6,45 |
8,75±0,46*** Cv=11,83 |
|
3-я опытная |
12,53±0,55 Cv=9,84 |
10,25±0,42* Cv=9,17 |
|
4-я опытная |
12,94±0,77 Cv=13,37 |
10,88±0,60 Cv=12,31 |
|
5-я опытная |
12,94±0,21 Cv=5,37 |
11,0±0,42 Cv=0,55 |
|
Контроль |
12,58±0,37 Cv=6,63 |
11,72±0,53 Cv=10,20 |
Таблица 31. Динамика уровня неконъюгированного билирубина у телят в летний период, мкмоль/л
| Группа |
До лечения |
После выздоровления |
|
1-я опытная |
11,85±0,52 Cv=9,72 |
7,81±1,54 Cv=43,99 |
|
2-я опытная |
9,66±1,21 Cv=27,95 |
8,45±0,54 Cv=14,34 |
|
3-я опытная |
9,61±2,56 Cv=59,65 |
9,46±1,22 Cv=28,9 |
|
4-я опытная |
9,1±3,08 Cv=75,81 |
8,94±0,53 Cv=13,17 |
|
5-я опытная |
10,87±0,5 Cv=10,37 |
9,83±0,89 Cv=20,31 |
|
Контроль |
8,72±1,35 Cv=34,66 |
8,82±1,66 Cv=42,11 |
Таблица 32. Динамика уровня неконъюгированного билирубина у телят в осенний период, мкмоль/л
| Группа |
До лечения |
После выздоровления |
|
1-я опытная |
10,71±0,54 Cv=11,33 |
10,96±1,57 Cv=32,05 |
|
2-я опытная |
10,48±1,08 Cv=23,14 |
9,58±0,9 Cv=21,05 |
|
3-я опытная |
10,22±0,45 Cv=9,77 |
10,32±1,12 Cv=24,15 |
|
4-я опытная |
9,89±0,68 Cv=15,35 |
10,78±0,38 Cv=7,90 |
|
5-я опытная |
10,02±0,64 Cv=14,30 |
10,7±0,36 Cv=7,60 |
|
Контроль |
10,33±0,28 Cv=5,98 |
11,07±0,47 Cv=9,42 |
При проведении исследований отмечены изменения в уровне аминотрансфераз. Содержание аланинаминотрансферазы (АЛТ) в сыворотке крови у телят 1–5-й опытных групп после выздоровления было ниже по сравнению с аналогичным показателем животных контрольной группы в летний период – на 25,9; 10,2; 9,8; 14,9; 20,9 %, осенью – на 21,7; 23,5; 18,1; 12,0; 7,1 % соответственно. По сравнению с исходными данными уровень АЛТ снизился у телят 1–5-й опытных и контрольной групп на 18,3; 14,2; 18,8; 22,5; 21,8 и 8,9 % – в летний, и на 23,0; 29,7; 24,8; 18,8; 16,0 и 8,1 % – в осенний периоды соответственно (табл. 33).
Уровень аспартатаминотрансферазы (АСТ) в сыворотке крови у телят 1–5-й опытных групп после выздоровления был ниже по сравнению с аналогичным показателем животных контрольной группы в летний период – в 2,5; 2,0; 2,5; 1,9; 2,3 раза, осенью – в 2,0; 2,4; 1,9; 2,2; 1,9 раз соответственно. По сравнению с исходными данными содержание АСТ у животных 1–5-й опытных групп увеличилось в 5,3; 4,0; 3,8; 1,9; 3,3 раза – в летний, и в 3,2; 4,2; 2,6; 5,0; 3,3 раза – в осенний периоды соответственно. В крови животных контрольной группы уровень АСТ по сравнению с исходными данными в летний период – уменьшился на 25,0 %, а в осенний период – увеличился на 33,3 % (табл. 34).
Таблица 33. Динамика уровня аминотрансфераз в крови телят в летний период, мкмоль/л
| Группа |
АЛТ |
АСТ |
||
|
До лечения |
После выздоровления |
До лечения |
После выздоровления |
|
|
1-я |
10,65±0,3 Cv=6,34 |
8,7±0,24 Cv=6,23 |
0,4±0,1 Cv=55,9 |
2,1±0,46 Cv=50,58 |
|
2-я |
12,3±0,35 Cv=6,33 |
10,55±0,2 Cv=4,24 |
0,45±0,09 Cv=46,48 |
1,8±0,36 Cv=45,43 |
|
3-я |
13,05±0,8 Cv=13,71 |
10,6±0,49 Cv=10,36 |
0,55±0,14 Cv=59,27 |
2,1±0,26 Cv=27,41 |
|
4-я |
12,9±1,31 Cv=22,63 |
10,0±0,26 Cv=5,86 |
0,9±0,41 Cv=101,3 |
1,75±0,14 Cv=15,31 |
|
5-я |
11,9±0,93 Cv=17,49 |
9,3±0,31 Cv=7,46 |
0,6±0,06 Cv=22,82 |
2,0±0,14 Cv=15,31 |
|
контроль |
12,9±0,56 Cv=9,75 |
11,75±0,4 Cv=7,52 |
0,65±0,23 Cv=79,76 |
0,6±0,1 Cv=37,27 |
Анализ хронобиологических особенностей влияния ветома 3 на уровень АЛТ и АСТ не позволяет сделать однозначных выводов. До лечения коэффициент де Ритиса был значительно понижен как в летний, так и в осенний периоды опыта. После выздоровления у телят опытных групп отмечена тенденция к повышению показателя данного диагностического теста, в основном за счет значительного повышения уровня аспартатаминотрансферазы, а также уменьшения аланинаминотрансферазы крови.
Таблица 34. Динамика уровня аминотрансфераз в крови телят в осенний период, мкмоль/л
| Группа |
АЛТ |
АСТ |
||
|
начало опыта |
окончание опыта |
начало опыта |
окончание опыта |
|
|
1-я |
11,5±0,37 Cv=7,21 |
8,85±0,39 Cv=9,91 |
0,5±0,11 Cv=50,0 |
1,6±0,13 Cv=17,82 |
|
2-я |
12,3±0,22 Cv=3,91 |
8,65±0,62 Cv=15,91 |
0,45±0,15 Cv=72,44 |
1,9±0,1 Cv=11,77 |
|
3-я |
12,3±0,35 Cv=6,33 |
9,25±0,31 Cv=7,4 |
0,6±0,15 Cv=55,9 |
1,55±0,12 Cv=17,67 |
|
4-я |
12,25±0,29 Cv=5,2 |
9,95±0,2 Cv=4,49 |
0,35±0,06 Cv=39,12 |
1,75±0,11 Cv=14,29 |
|
5-я |
12,5±0,33 Cv=5,83 |
10,5±0,21 Cv=4,45 |
0,45±0,15 Cv=72,44 |
1,5±0,22 Cv=33,33 |
|
контроль |
12,3±0,4 Cv=7,24 |
11,3±0,38 Cv=7,57 |
0,6±0,13 Cv=47,51 |
0,8±0,09 Cv=26,15 |
Рис. 33. Динамика коэффициента де Ритиса у телят в летний период
Коэффициент де Ритиса у телят 1–5-й опытных групп превышал после выздоровления аналогичный показатель животных контрольной группы в 4,6; 3,4; 3,9; 3,4; 4,2 раза – в летний, и в 2,5; 3,1; 2,3; 2,5; 2,0 раза – в осенний периоды соответственно (рис. 33).
В активной фазе действия ветома 3 на показатели коэффициента де Ритиса у телят в летний период находились значения 1-й и 5-й опытных групп, которые превышали мезор ((0,199±0,026) ед.) на 18,6 и 8,0 %, в пассивной фазе – 2, 3 и 4-й опытных групп, в которых коэффициент де Ритиса был ниже мезора на 14,1; 0,5 и 12,1 % соответственно. Абсолютная амплитуда показателей составила 0,07 ед., относительная амплитуда – 1,4 ед., коэффициент синхронизации – 1,4 ед./ч. Акрофаза влияния ветома 3 на коэффициент де Ритиса в летний период приходилась на 7 и 19 ч, что соответствует показателям телят 1-й опытной группы (рис. 33).
В активной фазе действия ветома 3 на показатели коэффициента де Ритиса у телят в осенний период находились значения 1-й и 2-й опытных групп, в которых данный показатель превышал мезор ((0,176±0,011) ед.) на 2,8 и 25,0 %, в пассивной фазе – показатели 3-й и 5-й опытных групп, в которых коэффициент де Ритиса был ниже мезора на 8,0 и 18,8 % соответственно, у телят 3-й опытной группы коэффициент де Ритиса соответствовал мезору. Абсолютная амплитуда показателей составила 0,08 ед., относительная амплитуда – 1,5 ед., коэффициент синхронизации – 0,51 ед./ч. Акрофаза влияния ветома 3 на коэффициент де Ритиса в осенний период приходилась на 8 и 20 ч, что соответствует показателям телят 2-й опытной группы (рис. 34).
Таким образом, при проведении исследований ветом 3 оказывал влияние на уровень аминотрансфераз у телят опытных групп, под действием препарата отмечена тенденция к повышению коэффициента де Ритиса, что может свидетельствовать об улучшении функционального состояния печени. Установлена зависимость действия пробиотика на уровень АЛТ, АСТ и коэффициента де Ритиса от времени применения препарата и сезона. Оптимальный эффект получен при применении ветома 3 в 7 и 19 ч – летом и в 8 и 20 ч – осенью.
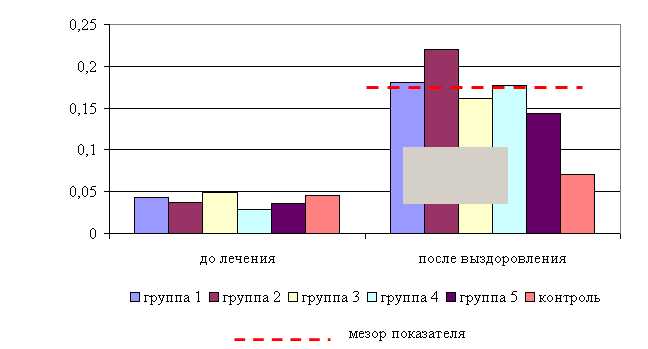
Рис. 34. Динамика коэффициента де Ритиса у телят в осенний период
Отмечено недостоверное влияние препарата на минеральный обмен веществ.
После выздоровления количество кальция в крови телят 1-й и 5-й опытных групп в летний период было выше по сравнению с аналогами из контроля 5,0 и 4,5 %, а у животных 2, 3 и 4-й опытных групп – ниже на 0,04; 0,3; 0,4 % соответственно; в осенний период уровень кальция в крови телят 1, 2 и 4-й опытных групп был выше аналогичного показателя телят контрольной группы на 1,4; 0,7 и 0,6 %, а у животных 3-й и 5-й опытных групп – ниже на 0,1 и 0,7% соответственно. По сравнению с исходными данными в летний период уровень кальция в крови телят 1, 2 и 5-й опытных и контрольной групп увеличился на 4,9; 1,5; 3,5 и 0,9 %, а у животных 3-й и 4-й опытных групп – уменьшился на 0,8 и 2,5 % соответственно. Количество кальция в осенний период увеличилось по сравнению с исходными данными у телят 1–5-й опытных групп на 4,4; 3,7; 0,1; 1,3; 0,2 % соответственно, тогда как у животных контрольной группы – уменьшилось на 0,64 % (табл. 36-37).
Уровень фосфора в крови телят 1–5-й групп после выздоровления был ниже аналогичного показателя животных контрольной группы на 6,3; 2,3; 2,3; 2,7; 4,8 % – в летний, и на 2,6; 5,1; 3,7; 3,6; 3,8 % – в осенний периоды соответственно. По сравнению с исходными данными уровень фосфора в летний период увеличился у телят 1-й и 2-й опытных и контрольной групп на 1,4; 0,3 и 0,3 %, а у животных 3, 4 и 5-й опытных групп – уменьшился на 3,0; 0,5 и 5,5 % соответственно. В осенний период количество кальция в крови телят 1–5-й опытных групп уменьшилось по сравнению с исходными данными на 2,9; 3,4; 1,0; 3,2; 2,7 % соответственно, а у аналогов из контроля – увеличилось на 0,8%. (табл. 35-36).
Таблица 36. Динамика показателей кальция и фосфора в крови телят летом, ммоль/л
| Группа |
Кальций |
Фосфор |
||
|
До лечения |
После выздоровления |
До лечения |
После выздоровления |
|
|
1-я |
2,662±0,048 Cv=4,08 |
2,792±0,05* Cv=4,01 |
1,234±0,052 Cv=9,49 |
1,251±0,026* Cv=4,69 |
|
2-я |
2,617±0,031 Cv=2,66 |
2,657±0,105 Cv=8,85 |
1,301±0,028 Cv=4,75 |
1,305±0,033 Cv=5,67 |
|
3-я |
2,673±0,037 Cv=3,13 |
2,651±0,059 Cv=4,95 |
1,345±0,035 Cv=5,78 |
1,305±0,025 Cv=4,36 |
|
4-я |
2,715±0,054 Cv=4,48 |
2,648±0,07 Cv=5,94 |
1,305±0,027 Cv=4,63 |
1,299±0,013 Cv=2,26 |
|
5-я |
2,684±0,041 Cv=3,39 |
2,778±0,042* Cv=3,41 |
1,345±0,04 Cv=6,72 |
1,271±0,047 Cv=8,27 |
|
Контроль |
2,683±0,058 Cv=4,87 |
2,658±0,038 Cv=3,16 |
1,311±0,027 Cv=4,57 |
1,335±0,029 Cv=4,89 |
Таблица 37. Динамика уровня кальция и фосфора в крови телят осенью, ммоль/л
| Группа |
Кальций |
Фосфор |
||
|
До лечения |
После выздоровления |
До лечения |
После выздоровления |
|
|
1-я |
2,545±0,018 Cv=1,61 |
2,657±0,014 Cv=1,2 |
1,366±0,018 Cv=2,94 |
1,327±0,027 Cv=4,51 |
|
2-я |
2,545±0,024 Cv=2,1 |
2,639±0,09 Cv=0,73 |
1,339±0,012 Cv=2,09 |
1,293±0,040 Cv=6,86 |
|
3-я |
2,616±0,020 Cv=1,73 |
2,619±0,031 Cv=2,64 |
1,328±0,011 Cv=1,86 |
1,313±0,034 Cv=5,82 |
|
4-я |
2,604±0,022 Cv=1,90 |
2,637±0,047 Cv=3,96 |
1,357±0,009 Cv=1,53 |
1,314±0,019 Cv=3,26 |
|
5-я |
2,596±0,024 Cv=2,08 |
2,602±0,024 Cv=2,1 |
1,347±0,009 Cv=1,53 |
1,311±0,025 Cv=4,34 |
|
Контроль |
2,638±0,025 Cv=2,12 |
2,621±0,022 Cv=1,91 |
1,352±0,016 Cv=2,72 |
1,363±0,014 Cv=2,34 |
Показатель кальций-фосфорного отношения по окончанию опыта в летний и осенний периоды изменился незначительно. Анализ сезонных изменений уровня кальция и фосфора в единице объема крови не позволяет сделать однозначных выводов.
Таким образом, ветом 3 обладает позитивным влиянием на белковый и пигментный обмен. Препарат нормализует функциональное состояние печени телят опытных групп, о чем свидетельствует увеличение коэффициента де Ритиса по сравнению с показателями контрольной группы в 4,6; 3,4; 3,9; 3,4; 4,2 раза – в летний, и в 2,5; 3,1; 2,3; 2,5; 2,0 раза – в осенний периоды соответственно, и снижение уровня небелковых азотистых компонентов крови. Оптимальный эффект получен при применении ветома 3 в 7 и 19 ч – летом, и в 8 и 20 ч – осенью.
3.4. Хронобиологические особенности действия ветома 3 на интенсивность роста телят после выздоровления
При изучении влияния циркадианных и сезонных биологических ритмов на интенсивность роста телят под влиянием ветома 3 установлено, что препарат обладает выраженным ростостимулирующим эффектом.
В летний период абсолютный прирост живой массы телят 1–5-й опытных групп был выше аналогичного показателя животных контрольной группы на 86,3; 51,0; 51,0; 39,2; 35,3 % соответственно (табл. 37).
Среднесуточный прирост живой массы телят 1–5-й опытных групп превышал в летний период аналогов из контроля 86,2; 50,9; 50,9; 39,1; 35,3 % соответственно (табл. 38).
Скорость роста телят 1–5-й опытных групп в летний период была выше по сравнению с животными контрольной группы на 86,2; 51,0; 51,0; 39,1; 35,3 % соответственно (рис. 36).
Таблица 37. Динамика абсолютного прироста живой массы телят, кг
| Группа |
Лето |
Осень |
|
1-я опытная |
19,0±2,77* Сv=32,66 |
19,0±1,76 Cv=20,72 |
|
2-я опытная |
15,4±2,73 Cv=39,66 |
21,2±2,13 Cv=22,47 |
|
3-я опытная |
15,4±3,25 Cv=47,18 |
19,0±1,41 Cv=16,64 |
|
4-я опытная |
14,2±1,43 Cv=22,49 |
17,0±2,7 Cv=35,54 |
|
5-я опытная |
13,8±2,15 Cv=34,93 |
17,4±2,01 Cv=25,89 |
|
Контроль |
10,2±3,44 Cv=75,43 |
15,4±3,31 Cv=48,07 |
Таблица 38. Динамика среднесуточного прироста живой массы телят, кг
| Группа |
Лето |
Осень |
|
1-я опытная |
0,633±0,92 Cv =32,61 |
0,636±0,059 Cv=20,85 |
|
2-я опытная |
0,513±0,091 Cv=39,68 |
0,706±0,071 Cv=22,5 |
|
3-я опытная |
0,513±0,108 Cv=47,18 |
0,633±0,047 Cv=16,65 |
|
4-я опытная |
0,473±0,48 Cv=22,51 |
0,566±0,09 Cv=35,55 |
|
5-я опытная |
0,460±0,072 Cv=35,05 |
0,58±0,067 Cv=25,89 |
|
Контроль |
0,340±0,115 Cv=75,44 |
0,513±0,110 Cv=48,07 |
В активной фазе влияния ветома 3 на уровень среднесуточных приростов живой массы телят в летний период находились показатели 1-й опытной группы, которые превышали мезор ((0,518±0,031) кг) на 22,2 %, в пассивной фазе – показатели 2, 3, 4 и 5-й опытных групп, которые были ниже мезора на 1,0; 1,0; 8,7 и 11,2 % соответственно. Продолжительность активной фазы превышала пассивную на 66,7 %. Абсолютная амплитуда показателей составила 0,17 кг, относительная амплитуда – 1,4 ед., коэффициент синхронизации – 0,34 ед./ч. Акрофаза влияния ветома 3 на уровень среднесуточного прироста живой массы телят в летний период приходилась на 7 и 19 ч, что соответствует показателям телят 1-й опытной группы (рис. 35).
В осенний период абсолютный прирост живой массы телят 1–5-й опытных групп был выше аналогичного показателя животных контрольной группы на 23,4; 23,4; 10,4; 13,0; 5,2 % соответственно (табл. 37).
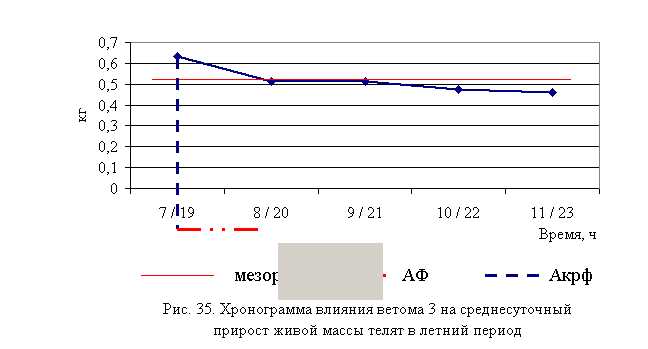
Среднесуточный прирост живой массы телят 1–5-й опытных групп превышал в летний период аналогов из контроля 23,9; 37,6; 23,4; 10,3; 13,1 % соответственно (табл. 38).
Скорость роста телят 1–5-й опытных групп в летний период была выше по сравнению с животными контрольной группы на 24,0; 37,6; 23,4; 10,3; 13,0 % соответственно (рис. 36).
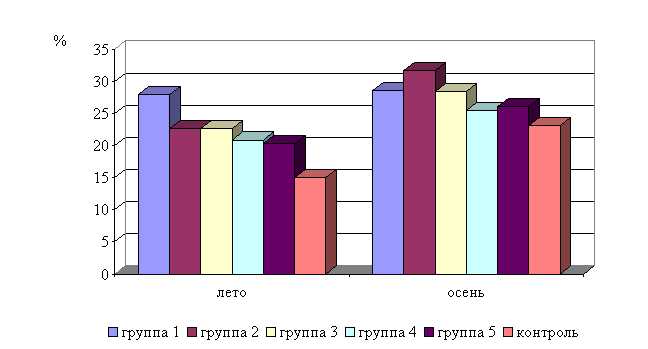
Рис. 36. Динамика скорости роста подопытных телят
В активной фазе влияния ветома 3 на среднесуточный прирост живой массы телят в осенний период находились показатели 1, 2 и 3-й опытных групп, которые превышали мезор ((0,624±0,025) кг) на 1,9; 13,1 и 1,4 %, в пассивной фазе – показатели 4 и 5-й опытных групп, которые были ниже мезора на 9,3 и 7,1 % соответственно. Продолжительность активной фазы превышала пассивную на 66,7 %. Абсолютная амплитуда показателей составила 0,14 кг, относительная амплитуда – 1,2 ед., коэффициент синхронизации – 0,6 ед./ч. Акрофаза влияния ветома 3 на среднесуточный прирост живой массы телят в осенний период приходилась на 8 и 20 ч, что соответствует показателям телят 2-й опытной группы (рис. 37).
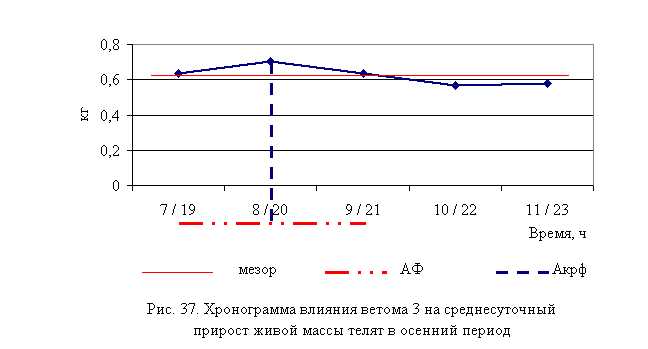
Таким образом, ветом 3 обладает выраженным ростостимулирующим эффектом. Установлена зависимость влияния пробиотика на среднесуточный прирост живой массы телят от времени применения препарата. Максимальное увеличение среднесуточного прироста живой массы и скорости роста отмечены у телят при применении ветома 3 в 7 и 19 ч – летом и 8 и 20 ч – осенью.
Анализ ростостимулирующего действия ветома 3 у телят показал выраженный эффект препарата на интенсивность роста животных. Оптимальная скорость роста телят была отмечена при применении препарата в 7 и 19 ч – в летний период, и в 8 и 20 ч – осенью, и превышала мезор показателя на 22,2 и 13,1 % соответственно. По сравнению с контролем у телят, получавших пробиотик по указанной схеме, скорость роста была выше на 86,2 и 37,6 % летом и осенью соответственно.
Увеличение скорости роста телят под влиянием пробиотика, по-видимому, связано как с купированием патологического процесса, а значит с повышением всасывающей способности кишечника, снижением потерь белка с экссулатом, так и со специфическим свойствами B. subtilis, которые продуцируют в организме 9--D-рибофуранозилгипоксантин, обладающий анаболическими свойствами (Беликов В.Г., 1993). Кроме того, сами бактерии, отмирая, становятся полноценным белковым субстратом для организма.






