2.1.2 Физиология спорообразующих бактерий
Способностью к образованию спор обладают аэробные, факультативные и анаэробные облигатные бактерии.
Первой анаэробной бактерией, открытой Луи Пастером в 1861 году, была Clostridium butyricum — спороносная палочка, вызывающая маслянокислое брожение углеводов. В дальнейшем было установлено, что спорообразующие и неспорообразующие облигатные анаэробы широко распространены в микробной экосистеме Земли. Анаэробы встречаются среди почти всех известных крупных систематических групп спорообразующих и неспорообразующих форм эубактерий, кокков, хламидобактерий, микобактерий и актиномицетов, вибрионов, спирилл и спирохет.
В настоящее время имеются три наиболее известные системы классификации анаэробов: Н. А. Красильникова (1949), Берги (1957) и Прево (1957, 1967). По Красильникову, все спорообразующие бактерии объединяются в семейство Васillасеае. Это семейство имеет два рода — Bacillus и Clostridium. В род Bacillus входят как аэробные, так и анаэробные бактерии (последние составляют Х-группу), которые не образуют гранулезы и не раздуваются (либо слабо раздуваются) при спорообразовании. Род Clostridium объединяет бактерии, клетки которых в процессе спорообразования раздуваются с приобретением клостридиальной (лимоновидной) формы и накапливают гранулезу.
Определитель Прево, Турпина и Кайзера наиболее полно описывает анаэробные микроорганизмы, известные в настоящее время микробиологам. Около 200 видов спорообразующих анаэробных бактерий распределяются в нем по трем порядкам: Clostridiales, Plectridiales и Sporovibrionales.
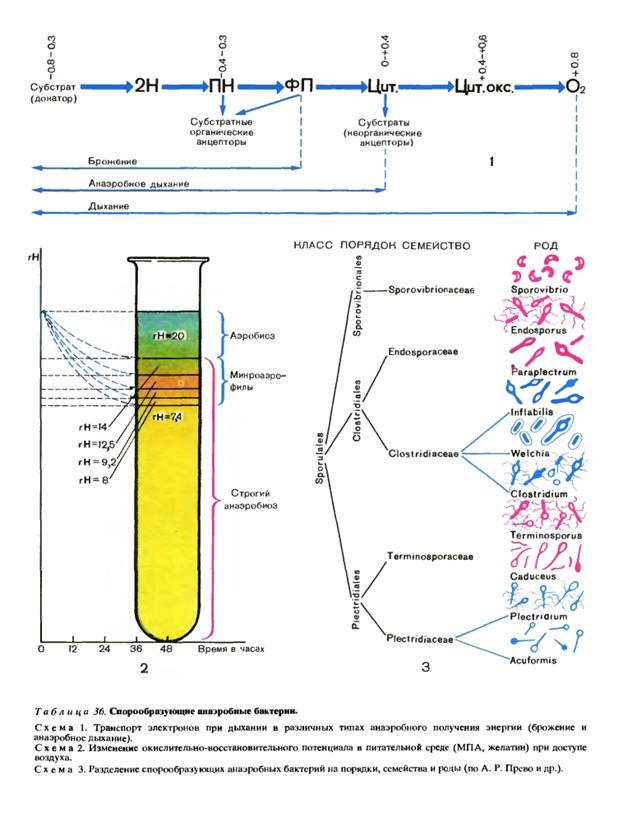
Морфологические различия между вегетативными клетками различных видов очень незначительны, а спорулирующие клетки и сами споры необычайно разнообразны. Это явление имеет очень большое значение для разделения анаэробов на систематические группы.
Для разделения на порядки служит форма вегетативных клеток (прямые или изогнутые палочки) и расположение спор в спорангиях (центральное, субтерминальное и терминальное). При разделении на семейства и роды принимаются во внимание окраска по Граму, подвижность клеток, способность образовывать капсулу. Различные сочетания этих признаков позволили разделить все спорообразующие анаэробные бактерии на 5 семейств и 10 родов.
Порядок Clostridiales. К нему относятся подвижные и неподвижные грамположительные и грамотрицательные палочки, раздувающиеся при спорообразовании. Споры расположены центрально и субтерминально. Имеются пигментированные виды. В основном анаэробные, но возможны факультативные формы. В порядок входят два семейства.
Семейство Endosporaceae включает грамотрицательные клостридии. Споры в них расположены центрально или субтерминально. В этом семействе два рода: Endosporus - подвижные формы и Paraplectrum - неподвижные. Оба образуют капсулу.
Семейство Clostridiaceae включает грамположительные клостридии. Споры центральные или субтерминальные. Подразделяется на три рода: Inflabilis - неподвижные формы, Welchia - неподвижные капсулированные палочки, Clostridium - подвижные формы.
Порядок Plectridiales. Подвижные и неподвижные, грамположительные и грамотрицательные палочки. Споры образуются терминально. Порядок подразделяется на два семейства.
Семейство Terminosporaceae - грамотрицательные плектридии. Семейство состоит из двух родов: Terminosporus — подвижные и Caduceus — неподвижные формы.
Семейство Plectridiaceae - грамположительные плектридии. В этом семействе два рода: Plectridium - подвижные и Acuformis - неподвижные организмы.
Порядок Sporovibrionales включает спорообразующие подвижные анаэробные бактерии с изогнутыми (вибрионоподобными) грамотрицательными клетками. Большинство из них способно восстанавливать сульфаты и нитраты.
В этом порядке одно семейство - Sporovibrionaceae. В него входит один род - Sporovibrio, объединяющий четыре вида спорообразующих анаэробов. Один из этих видов - Sporovibrio ferrooxydans. Он не восстанавливает сульфаты, а железо окисляет в гидрат железа.
По Кэмпбеллу и Постгейту, все спорообразующие анаэробы, обладающие постоянной способностью восстанавливать сульфаты, были выделены в новый род - Desulfotomaculum. В него входят облигатные анаэробы с грамотрицательными, прямыми или изогнутыми палочками, раздувающимися у термофильных форм. Споры формируются терминально или субтерминально. Бактерии подвижные1, движение перитрихиальное1, восстанавливают сульфаты до сульфидов. В их составе присутствуют цитохромы2 (гемопротеины3). Состав ДНК колеблется в пределах 41,7—49,2 мол.% Г+Ц.
Неспоровые сульфатредуцирующие бактерии4 из рода Desulfovibrio четко отличаются от спорообразующих по проценту Г+Ц в ДНК, наличию пигмента десульфовиридина и цитохрома, монотрихиальным жгутикованием и отсутствием спорообразования.
Род Desulfotomaculum включает четыре вида. Desulfotomaculum nigrificans - типовой вид. Споры овоидные, субтерминальные. Бактерии могут расти без сульфатов на среде с пируватом. Глюкозу и другие углеводы не используют, термофильные. Температурный оптимум 55 °С. Процент Г+Ц в ДНК - 49,2.
Desulfotomaculum ruminis. Споры овоидные, терминальные или субтерминальные. Бактерии способны расти без сульфатов на среде с пируватом. Глюкозу и другие углеводы не сбраживают, мезофилы, Г+Ц7 в ДНК - 49,2 %.
Desulfotomaculum orientis. Споры круглые, могут располагаться в клетках центрально, эксцентрально и терминально. Глюкозу и другие углеводы бактерии не сбраживают, не растут на средах с пируватом без сульфатов, мезофилы. Процент Г+Ц7 в ДНК - 45,1.
Японские авторы описали еще один вид спорообразующих сульфатредуцирующих бактерий - Desulfotomaculum antarcticum. Споры этих бактерий овоидные, располагаются в клетках центрально или терминально. Нуклеотидный состав ДНК неизвестен. Этот вид отличается от других видов рода Desulfotomaculum способностью использовать глюкозу и разжижать желатин.
Спорообразующие облигатные анаэробы
Спорообразующие облигатные анаэробы изучать достаточно сложно, поскольку они растут в условиях практически полного отсутствия кислорода, что требует наличия специального оборудования обеспечивающего поддержание требуемого состава газовой среды и температурного режима в процессе выполнения опыта. В настоящее время наиболее распространённым прибором для проведения культуральных работ с облигатными анаэробами является анаэростат, который достаточно прост и удобен в работе и позволяет создать и поддерживать в процессе эксперимента требуемые параметры газовой среды и температуры. И, тем не менее, широкомасштабные опыты с использованием анаэростатов ставить затруднительно. В этой связи количество изученных облигатных анаэробов, в том числе и спорообразующих, в настоящее время невелико. Тем более не велико количество облигатных анаэробов, для которых выделена чистая культура и детально изучены их свойства в области физиологии, цитологии, генетики микроорганизма.
Факультативные анаэробы
Факультативные анаэробы, способны жить как за счет дыхания, используя молекулярный кислород, так и за счет «нитратного дыхания», либо брожения различных органических веществ в анаэробных условиях.
По современным представлениям, аэробные спорообразующие бактерии, или бациллы5, объединяются в отдельный род Bacillus семейства Bacillaceae. Этот род, включающий много разнообразных видов, имеет ряд характерных особенностей и отличается от других бактериальных организмов комплексом морфолого-физиологических признаков, из которых наиболее важными являются палочковидная форма клеток, способность образовывать эндоспоры, потребность в свободном кислороде для роста.
Общая краткая характеристика объединяемых в род бацилл аэробных спорообразующих бактерий сводится к следующему.
Бациллы - свободноживущие, одноклеточные, нефотосинтезирующие, аэробные, палочковидные клетки, образующие типичные эндоспоры. Относятся к гетеротрофным6 организмам. Размножаются поперечным делением клеток. Ветвление и почкование клеток как способ размножения не отмечены. Поперечный размер клеток варьирует в пределах 0,4-2 мкм. Вегетативные клетки имеют вид прямых или слабоизогнутых палочек с параллельными сторонами и округлыми концами, которые в редких случаях резко обрезанные.
Расположение клеток различное - от одиночных до длинных цепочек. За исключением сибиреязвенной бациллы, бактерии этого рода подвижные, снабжены перитрихиально1 расположенными жгутиками.
Большинство видов является грамположительными бактериями, часть - грамвариабильными. Клетки хорошо окрашиваются обычными анилиновыми красками. Ни один представитель рода не является типично кислотоустойчивым организмом. У многих видов отмечается наличие внутриклеточного жира, гликогена9, волютина10 и других включений. Капсула встречается лишь у сибиреязвенной бациллы и некоторых других видов при специфических условиях роста.
В бактериальных клетках в общем количестве оснований ДНК 32-65 мол. % гуанина и цитозина.
Большинство видов, исключая некоторые, главным образом энтомопатогенные11, формы, хорошо растет на мясопептонном агаре (МПА) при реакции среды, близкой к нейтральной. Отдельные виды развиваются в щелочной среде и требуют особых источников азота или углерода.
Культуральные особенности видов, выросших на разных средах, резко различны. На твердых питательных средах образуются колонии от 1-2 до 5 мм и более в диаметре: гладкие, зернистые, пленчатые, складчато-морщинистые и сухие, слизеобразующие и пастообразные с характерной структурой края. При развитии на жидких средах обнаруживается тенденция к образованию поверхностной пленки.
Встречаются виды, образующие на поверхности агаризованных сред подвижные колонии.
Большинство видов бактерий активно продуцируют ферменты, гидролизующие белки, крахмал и другие субстраты. Многие виды обладают антагонистическими свойствами и вырабатывают антибиотики полипептидной природы8. Отдельные виды нуждаются в присутствии витаминов, аминокислот и других дополнительных факторов роста. Кислоту и газ продуцируют лишь бактерии некоторых видов, все остальные при росте на углеводах образуют одну кислоту. Оптимальная температура роста обычно варьирует в пределах 30—40° С. Встречаются виды, развивающиеся при температуре ниже 12 °С и выше 50 °С.
Некоторые аэробные бактерии — возбудители болезней. Вас. anthracis вызывает сибирскую язву у человека и животных; Вас. larvae — возбудитель американского гнильца медоносной пчелы; Вас. alvei и Вас. pulvifaciens трактуются как организмы, играющие определенную роль в болезнях пчел; Вас. popilliae и Вас. lentimorbus — возбудители молочной болезни японского жука; некоторые виды группы Вас. cereus-thuringiensis вырабатывают специфические энтомоцидные токсины.
Классификация видов аэробных спорообразующих бактерий разработана недостаточно полно. Одной из причин этого является ограниченность различий во внешних признаках бактерий. Известно, что большинство видов различаются между собой малозначимыми признаками строения и развития клеток, по форме колоний, а также физиологическим признакам. Многие вопросы биологии спороносных бактерий требуют дополнительных глубоких исследований.
Хорошо изучены только те виды, которые имеют практическое значение (патогенные, бродильные формы, продуценты различных антибиотиков, ферментов и т. д.). Вопросы систематики спорообразующих бактерий изучались специалистами прикладных наук для диагностических целей, в основном санитарно-гигиенического назначения.
Для выделения бактерий в самостоятельные виды одни авторы считают решающими ферментативные свойства организмов, другие — морфологические и культуральные признаки, третьи — цитологические особенности бактериальной клетки и т. д. Во всех подобных случаях принципы систематики меняются по мере выявления новых особенностей в строении и физиологических свойствах организмов и использования их в качестве признаков видовой идентификации. Систематика разных видов спорообразующих бактерий отражает степень их изученности, а следовательно, должна развиваться и изменяться по мере накопления новых данных.
Вопросы систематики бактерий представляют исключительную важность для работ по изысканию и изучению образования разнообразных физиологически активных веществ микробного происхождения. Биологический вид представляет единство специфических морфологических и физиолого-биохимических признаков организма, определяющих все особенности его жизнедеятельности, распространения и взаимодействия с внешней средой. В связи с этим образование того или иного продукта жизнедеятельности, как и характерные биологические особенности, невозможно представить в отрыве от видовой принадлежности микроорганизма.
Работы в области систематики спорообразующих бактерий и определения их положения в мире микроорганизмов начались еще до открытия спор у бактерий. Термин Bacillus на разных этапах развития микробиологии использовался вольно для обозначения как спороносных, так и неспороносных бактерий. В сущности, подобное положение сохранилось до наших дней, и мы нередко затрудняемся не только идентифицировать описанные виды, но и установить их истинную принадлежность к спорообразующим бактериям. Многие спорообразующие бактерии именовались различными авторами под родовыми названиями, которые в настоящее время служат обозначением родов неспороносных бактерий и других микроорганизмов, что еще более осложняет точное представление об описываемом организме.
Таксономия12 бактерий раннего периода в основном была конструктивной и имела диагностическое назначение. На этом этапе рассматривались подходы к созданию крупных систематических категорий.
По мере накопления фактического материала начали предпринимать попытки классификации по аналитическому принципу, учитывая родственные взаимоотношения микроорганизмов.
Систематика спорообразующих бактерий развивалась по мере эволюции взглядов на принципы дифференциации и идентификации микробных видов, а также усовершенствования методов морфолого-фиэиологического изучения бактерий.
Наиболее ранними в этом отношении были попытки микробиологов разграничить виды спорообразующих бактерий по морфологическим признакам, в частности по форме спор, способу их прорастания и т. п. Малозначимость этих признаков дифференциации и их неспецифичность привели к тому, что в дополнение к предыдущим описаниям было идентифицировано много новых видов, большинство из которых в последующем не были приняты или трактуются сейчас как синонимы других систематических категорий.
Широкое распространение спорообразующих бактерий в окружающей среде и их частое обнаружение в различных продуктах, особенно при их порче, вызвали к ним определенный интерес. Первые работы по классификации этой группы бактерий были выполнены в основном представителями пищевой и санитарной микробиологии. При этом одни авторы основывались на морфологических признаках микроорганизмов, а другие — на физиологических.
Классификация, основанная на различиях в способности аэробных спорообразующих бактерий сбраживать субстраты, предусматривала подразделение этих организмов на три группы по способности сбраживать глюкозу и образовывать ацетилметилкарбинол. Согласно другой классификации, которую в настоящее время используют наиболее часто, данный род бактерий разделяется на три группы по соотношению поперечных размеров спор и вегетативных клеток.
Группа I — наиболее обширная — включает виды спороносных бактерий, у которых не отмечается отчетливого раздувания спорангия13 в процессе образования спор.
Группа II объединяет бактерии, образующие овальные споры, раздувающие спорангий13.
Группа III охватывает бациллярные виды, характеризующиеся округлыми или шаровидными спорами, раздувающими спорангий13; объединяет редко встречающиеся виды спорообразующих бактерий.
Вопросы классификации различных видов аэробных спорообразующих бактерий разрабатывались многими отечественными авторами. Особенно тщательно были изучены морфологофизиологические особенности отдельных групп и видов этих бактерий Е. Н. Мишустиным с сотрудниками. Данные этих исследований свидетельствуют о большом многообразии видов и экологических разновидностей спорообразующих бактерий в зависимости от мест их обитания, почвенно-климатической зональности и микробного ценоза разных типов почв.
В настоящее время для определения видов спорообразующих бактерий наряду с морфолого-физиологическими особенностями используют и многие другие признаки.
Важными критериями для определения и дифференциации бактерий являются отношение к действию специфичных фагов, агглютинация с гомологичными сыворотками к споровому, соматическому и жгутиковому антигенам, рост при высокой концентрации солей, различной температуре и т. п.
Многие отмеченные особенности применяются для внутривидового подразделения и выделения культур спороносных бактерий в отдельные разновидности, серотипы и другие мелкие систематические подразделения. В ряде случаев некоторые признаки служат основой для выделения культур спорообразующих бактерий в новые виды. Так, например, в качестве новых видов описаны психрофильные14, развивающиеся при низкой температуре культуры спорообразующих бактерий. Подобным же образом выделены некоторые активные продуценты амилолитических и целлюлолитических ферментов. В литературе было описано много новых видов бацилл, патогенных для некоторых насекомых, растений и животных, что в большинстве случаев не подтвердилось в последующие годы.
Успехи в области создания счетно-вычислительной техники дали основу для быстрой математической обработки данных по родству и отдаленности организмов. Применение этой техники в бактериальной таксономии12 получило название числовой таксономии и было введено в микробиологию в последние годы. Числовая таксономия рассматривает группирование и идентификацию организмов с учетом их подобия при анализе не менее чем 50—60 объективных признаков. Этот метод с разработкой схем подобия и разграничения уже использован рядом авторов для систематики различных видов спороносных бактерий.
Особого внимания заслуживает разработка новых подходов к трактовке вопросов филогенетической15 таксономии бактерий с помощью биохимических исследований. Наибольший интерес в этом отношении представляют попытки классификации бактерий с использованием новейших данных молекулярной биологии.
Аэробные бактерии имеют гамму различных физиологических признаков.Наличие у споровых бактерий разных механизмов переноса электронов обусловливает отличие их физиологических признаков при культивировании в различных условиях. Так, клетки Bac. subtilis при выращивании на сложной триптон - дрожжевой среде с глюкозой характеризуются активным дыханием и одновременно снижением способности к ферментации, а клетки, выращенные на синтетической среде, теряют способность к брожению и окисляют глюкозу в процессе дыхания. При этом выявлены качественные изменения в составе ферментов.
По результатам исследований Головачева и соавт., (1965), Логинова и соавт. (1973), большинство аэробных бацилл являются мезофилами и активно развиваются при 30-45 0С, но встречаются и термофилы, развивающиеся при 65 0С и выше. На Камчатке из воды горячих источников выделены Bac. stearothermophilus, Bac. brevis, Bac. subtilis, Bac. cereus, Bac. megaterium, Bac. licheniformis, которые активно развиваются при температуре в пределах 65 0С (Логинова и др., 1976). Установлено, что отдельные штаммы Bac. subtilis, Bac. licheniformis, Bac. cereus размножаются при 7 0С и ниже (Tinuone, Harmon, 1975). Эти аэробные спорообразующие бактерии являются психрофильными и обнаружены в Антарктике (Nelson, Parcinson, 1978).
Для большинства бацилл оптимальной для развития является рН 7 с колебаниями от 2 до 8. Bac. alcolophilus активно развивается при рН 9-10, а Bac. aсidocaldarius при 3-4 (Bergeys, 1974).
Концентрацию солей в питательной среде бациллы могут переносить от 2 до 25 %, а отдельные штаммы Bac. brevis, Bac. cereus, Bac. circulans от 5 до 30 % (Тимук, 1979).
Термофильные бациллы одинаково хорошо растут как в аэробных, так и в анаэробных условиях. Это объясняется наличием у термофилов более активных анаэробных дегидрогеназ (Позмогова, Мальян, 1976).
Согласно исследованиям Wangetal. (1962), у бактерий рода Bacillus преобладает гликолитический16 путь расщепления углеводов независимо от наличия в среде кислорода.
Бациллы для окисления используют простые (моно- и дисахариды) и сложные высокомолекулярные (пектин) углеводы, полисахариды (декстрин, крахмал), органические кислоты и спирты.
Бактерии рода Bacillus обладают способностью интенсивно окислять углеводороды - парафин, гексодекан, a-олефины ( Тархова, 1973). При температуре инкубации 37 0С клетки Bac. subtilis при потреблении 1 моля глюкозы образуют 4 моля АТФ. С повышением температуры и концентрации глюкозы процесс образования АТФ замедляется и это, по-видимому, связано с тем, что гликолиз при повышенной температуре переключается на цикл Энтнера-Дудорова17 (Ibraghim, 1973).
Бациллы обладают свойством аммонификации18 белковых соединений. Протеолитические ферменты19 бацилл вызывают гидролиз пептидных связей в молекулах белка с образованием пептидов и олигопептидов, которые под действием ферментов расщепляются до свободных аминокислот, либо используются для синтеза клеточного белка. Бациллы могут выборочно утилизировать некоторые аминокислоты. Так, Bac. subtilis проявляет хемотаксис20 в отношении l-форм аминокислот, в присутствии b-форм аминокислот хемотаксической реакции не обнаружено (Drift, 1974).
Бациллы обладают свойством усваивать азот. Причем одни лучше усваивают аммонийный азот, другие - азот нитратов. Например, Bac. subtilis- mesentericus легко усваивают аммонийные соединения. Для бацилл в качестве азотного питания целесообразнее использовать пептон, гидролизат казеина, автолизат дрожжей.
Бациллы также обладают свойством усваивать минеральные соединения азота (аммонийные соли, нитриты) и могут развиваться и на белковой, и на синтетической среде с нитритами. Они восстанавливают нитриты и переводят их в органические азотсодержащие соединения. Наиболее активно восстанавливают нитриты Bac. licheniformis и Bac. cereus.
Микроэлементы также необходимы для нормального роста и развития бацилл. Согласно исследованиям Beveridge, Murray (1976), клеточные стенки являются барьером только для ионов Li+, Ba+, Co2+и Al3+. Наличие в питательной среде необходимых ионов металлов активизирует накопление биомассы и повышает гидролитическую активность клеток. При недостатке магния, фосфора и других элементов происходит замедление роста вегетативных клеток Bac. stearothermophilus (Lee, Brrown,1975). Биосинтез протеаз Bac. subtilis активизирует Mg2+, а Cu2+ и Zn2+ подавляют их биосинтез (Naumski, 1973). Ионы Са стимулируют амилолитическую активность Bac. subtilis, выраженность этого процесса зависит от соотношения ионов Са и Mg(Кюдулене и соавт., 1975). Ионы К стимулируют синтез b-амилазы Bac.cereus (Shinkeetal., 1977). Ионы Mn2+ участвуют в процессе спорообразования у Bac. subtilis, Bac. cereus, Bac. megaterium в качестве кофактора фосфатглицеромутазы, которая необходима для процесса споруляции (Vasantha, Freese, 1979).
Рост бацилл зависит от концентрации ионов металлов в среде. С увеличением концентрации ионов металлов их рост замедляется. Однако выделены бациллы, обладающие высокой устойчивостью к повышенному содержанию в почве Zn, Cu, Mg (Летунова, Белова,1978). Устойчивость бактерий к ртути у многих штаммов рода Bacillus связана с их способностью восстанавливать соединения ртути.
Бациллы испытывают большие потребности в фосфоре и многие из них способны переводить нерастворимые соединения фосфора в растворимые. Например, Bac. polymyxa и Bac. megaterium способны растворять Ca3(PO4)2 и разные фосфаты. Ряд бацилл могут анаэробно усваивать фосфит и гипофосфит на глюкозо - минеральной среде. Спорообразующие анаэробные бактерии чаще минерализуют органофосфаты, что используется для получения фосфобактерина.
Сенная палочка обладает способностью образовывать пигмент на среде с углеводами независимо от наличия тирозина в среде. По данным В.В. Смирнова и соавт. (1982), пигмент можно отнести к меланинам. Пигменты, синтезируемые Bac. subtilisv. niger 16 k, окисляются под действием H2O2 и KMnO4. Аскорбиновая кислота является ингибитором меланиногенеза (Shiehetal., 1974). Установлено, что на синтетической среде с глюкозой и со смесью аминокислот, взятых в качестве единственного источника азота (аспарагин, лейцин, валин, триптофан, тирозин, фенилаланин), исследуемые бактериальные культуры продуцируют пигмент черного цвета через 72 ч; при уменьшении концентрации аминокислот с 0,5 до 0,25 % образуется грязно- бурый пигмент, а при исключении комплекса микроэлементов колонии остаются белыми. При удалении же из среды аланина, аспарагина и валина вообще не наблюдается пигментообразование. Таким образом, существенное влияние, которое оказывают различные аминокислоты на процесс образования и характер пигмента, свидетельствует о разнообразии механизмов синтеза этих соединений.
Выявлено, что в процессе образования меланина микроорганизмами на определенной стадии синтезируется диоксифенилаланин (ДОФА), который представляет собой ароматическую аминокислоту, выполняющую в организме важную роль, участвуя в образовании биологически активных соединений, в частности адреналина.
Химическая природа этих пигментов окончательно не выяснена, хотя они представляют значительный интерес, поскольку с ними связывают защитные свойства клеток. В кислотном гидролизате полисахарида термофильной культуры Bac. subtilis 20 обнаружены ксилоза, манноза и глюкоза (Сергеев, Ложнова, 1975).
В подавляющем большинстве случаев бациллы образуют каталазу.
Сенная палочка разлагает ксилон (гемицеллюлозу), который по распространению в природе занимает второе место после целлюлозы. Bac. subtilis, Bac. mesentericus усваивают лигнин и целлюлозу (Bellamy, 1973; Tewari, Chahal, 1977). Термофильные бактерии более активны.
Таким образом, для нормального роста и развития бактерий необходимы микро- и макроэлементы, и, изменяя их концентрацию в среде, можно ускорить или замедлить рост бацилл. Наличие различных путей усвоения углерода, широкие ассимиляционные возможности способствуют распространению этих бактерий в природе и обусловливают их адаптивные свойства. Большое практическое значение имеет свойство ряда бацилл восстанавливать нитриты, расщеплять ксилон и усваивать лигнин и целлюлозу.






